
Variable Fitness Response of Two Rotifer Species Exposed to Microplastics Particles: The Role of Food Quantity and Quality


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of Organisms
2.2. Microplastics
2.3. Experimental Procedure
2.4. Statistical Analysis
3. Results
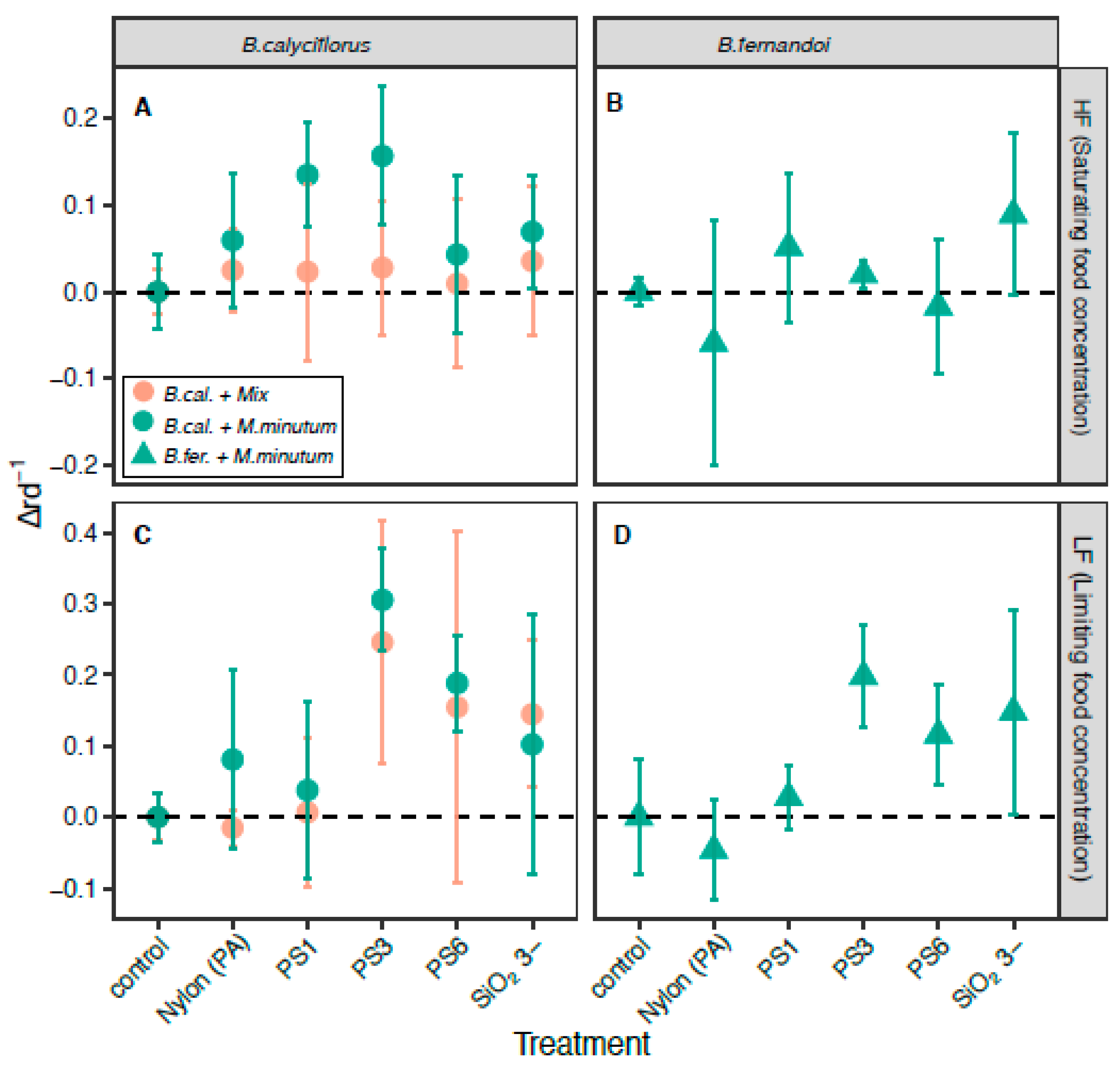
3.1. Effect of the MP Beads on Population Growth Rate
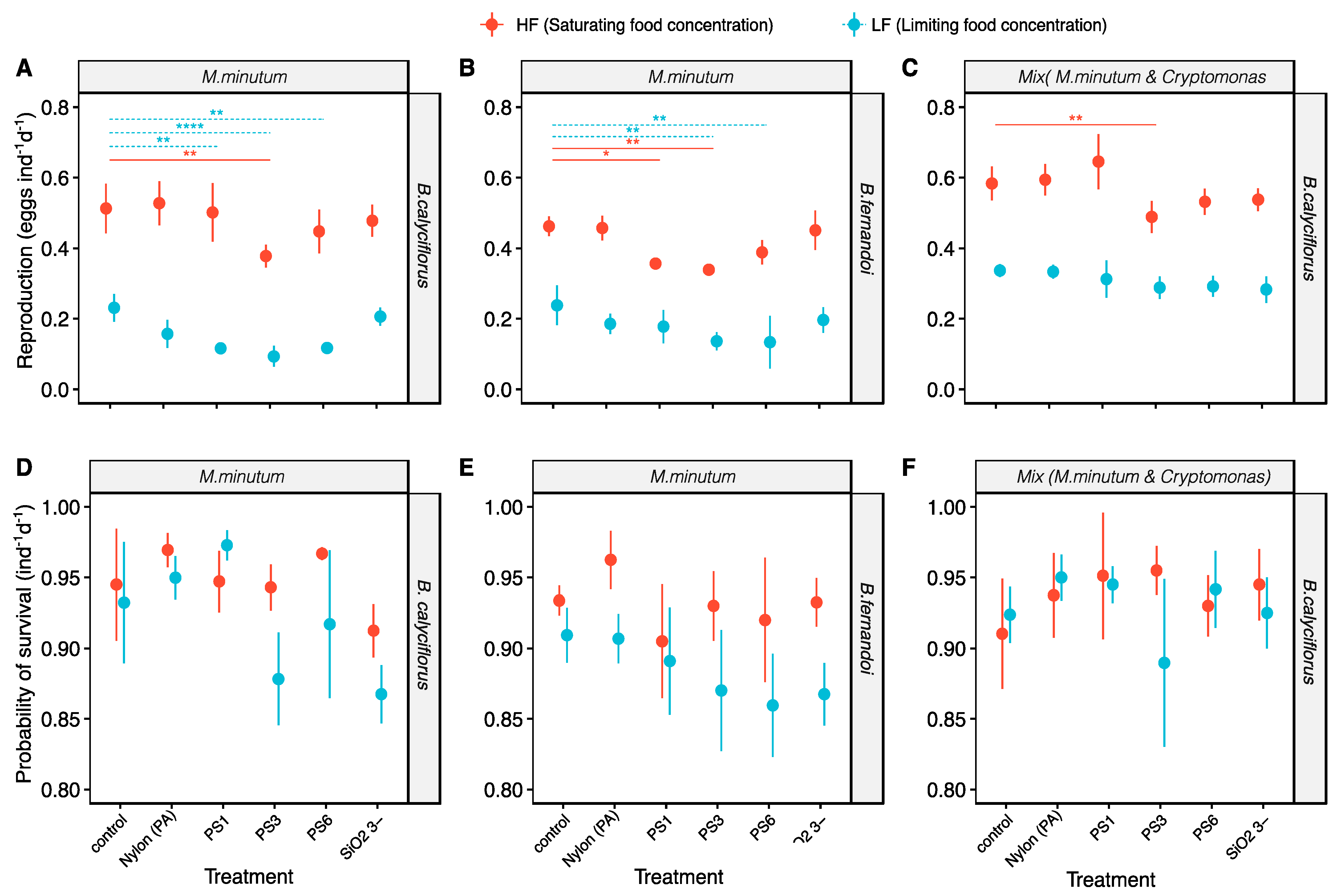
3.2. Effect of the MP Beads on Reproduction
3.3. Effect of the MP Beads on Survival
4. Discussion
4.1. The Role of Food Quantity and Food Quality on Microplastics Effect
4.2. Size Particles Effect
4.3. Silica and (PA) Nylon Microbeads
4.4. Ecological Relevance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, C.J. Synthetic polymers in the marine environment: A rapidly increasing, long-term threat. Environ. Res. 2008, 108, 131–139. [Google Scholar] [CrossRef]
- Eerkes-Medrano, D.; Thompson, R.C.; Aldridge, D.C. Microplastics in freshwater systems: A review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water Res. 2015, 75, 63–82. [Google Scholar] [CrossRef]
- Li, J. Microplastics in freshwater systems: A review on occurrence, environmental effects, and methods for microplastics detection. Water Res. 2018, 137, 362–374. [Google Scholar] [CrossRef]
- Murphy, F.; Ewins, C.; Carbonnier, F.; Quinn, B. Wastewater treatment works (WwTW) as a source of microplastics in the aquatic environment. Environ. Sci. Technol. 2016, 50, 5800–5808. [Google Scholar] [CrossRef] [Green Version]
- Kole, P.J.; Löhr, A.J.; Van Belleghem, F.G.; Ragas, A.M. Wear and Tear of Tyres: A Stealthy Source of Microplastics in the Environment. Int. J. Environ. Res. Public Health 2017, 14, 1265. [Google Scholar] [CrossRef]
- Corradini, F.; Meza, P.; Eguiluz, R.; Casado, F.; Huerta-Lwanga, E.; Geissen, V. Evidence of Microplastic Accumulation in Agricultural Soils from Sewage Sludge Disposal. Sci. Total Environ. 2019, 671, 411–420. [Google Scholar] [CrossRef]
- Liu, K.; Wang, X.; Wei, N.; Song, Z.; Li, D. Accurate Quantification and Transport Estimation of Suspended Atmospheric Microplastics in Megacities: Implications for Human Health. Environ. Int. 2019, 132, 105127. [Google Scholar] [CrossRef]
- Loppi, S.; Roblin, B.; Paoli, L.; Aherne, J. Accumulation of Airborne Microplastics in Lichens from a Landfill Dumping Site (Italy). Sci. Rep. 2021, 11, 4564. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.; Webb, H.; Lindeque, P.K.; Fileman, E.S.; Halsband, C.; Galloway, T.S. Isolation of microplastics in biota-rich seawater samples and marine organisms. Sci. Rep. 2015, 4, 4528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Sá, L.C.; Oliveira, M.; Ribeiro, F.; Rocha, T.L.; Futter, M.N. Studies of the Effects of Microplastics on Aquatic Organisms: What Do We Know and Where Should We Focus Our Efforts in the Future? Sci. Total Environ. 2018, 645, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Bermúdez, J.R.; Metian, M.; Oberhänsli, F.; Taylor, A.; Swarzenski, P.W. Preferential Grazing and Repackaging of Small Polyethylene Microplastic Particles (≤5 Μm) by the Ciliate Sterkiella sp. Mar. Environ. Res. 2021, 166, 105260. [Google Scholar] [CrossRef]
- De Ruijter, V.N.; Redondo-Hasselerharm, P.E.; Gouin, T.; Koelmans, A.A. Quality Criteria for Microplastic Effect Studies in the Context of Risk Assessment: A Critical Review. Environ. Sci. Technol. 2020, 54, 11692–11705. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Xu, W.; Gu, Q.; Chen, Y.; Zhou, Q.; Zhang, L.; Yang, Z. Small-Sized Microplastics Negatively Affect Rotifers: Changes in the Key Life-History Traits and Rotifer–Phaeocystis Population Dynamics. Environ. Sci. Technol. 2019, 53, 9241–9251. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.-H.; Sun, Z.-X.; Feng, L.-S.; Jin, T.; Xing, J.-C.; Wen, X.-L. Algal Density Affects the Influences of Polyethylene Microplastics on the Freshwater Rotifer Brachionus Calyciflorus. Chemosphere 2021, 270, 128613. [Google Scholar] [CrossRef] [PubMed]
- Canniff, P.M.; Hoang, T.C. Microplastic Ingestion by Daphnia Magna and Its Enhancement on Algal Growth. Sci. Total Environ. 2018, 633, 500–507. [Google Scholar] [CrossRef] [Green Version]
- Beiras, R.; Bellas, J.; Cachot, J.; Cormier, B.; Cousin, X.; Engwall, M.; Gambardella, C.; Garaventa, F.; Keiter, S.; Le Bihanic, F.; et al. Ingestion and Contact with Polyethylene Microplastics Does Not Cause Acute Toxicity on Marine Zooplankton. J. Hazard. Mater. 2018, 360, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Christensen, N.D.; Wisinger, C.E.; Maynard, L.A.; Chauhan, N.; Schubert, J.T.; Czuba, J.A.; Barone, J.R. Transport and Characterization of Microplastics in Inland Waterways. J. Water Process Eng. 2020, 38, 101640. [Google Scholar] [CrossRef]
- Kruse, J.; Laermanns, H.; Stock, F.; Foeldi, C.; Schaefer, D.; Scherer, C.; Bogner, C. Proceedings of the Microplastic in fluvial environments - an example of the Elbe river near Dessau-Roßlau, Germany, EGU General Assembly 2021, online, 19–30 April 2021. EGU21-2686. [CrossRef]
- Free, C.M.; Jensen, O.P.; Mason, S.A.; Eriksen, M.; Williamson, N.J.; Boldgiv, B. High-Levels of Microplastic Pollution in a Large, Remote, Mountain Lake. Mar. Pollut. Bull. 2014, 85, 156–163. [Google Scholar] [CrossRef]
- Frydkjær, C.K.; Iversen, N.; Roslev, P. Ingestion and Egestion of Microplastics by the Cladoceran Daphnia Magna: Effects of Regular and Irregular Shaped Plastic and Sorbed Phenanthrene. Bull. Environ. Contam. Toxicol. 2017, 99, 655–661. [Google Scholar] [CrossRef]
- Merriman, J.L.; Kirk, K.L. Temporal patterns of resource limitation in natural populations of rotifers. Ecology 2000, 81, 141–149. [Google Scholar] [CrossRef]
- Cordova, S.E.; Giffin, J.; Kirk, K.L. Food Limitation of Planktonic Rotifers: Field Experiments in Two Mountain Ponds: Food Limitation. Freshw. Biol. 2001, 46, 1519–1527. [Google Scholar] [CrossRef]
- Weithoff, G. Vertical Niche Separation of Two Consumers (Rotatoria) in an Extreme Habitat. Oecologia 2004, 139, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Mayagoitia, E.; Ciros-Perez, J.; Sanchez-Martinez, M. A Story of Famine in the Pelagic Realm: Temporal and Spatial Patterns of Food Limitation in Rotifers from an Oligotrophic Tropical Lake. J. Plankton Res. 2011, 33, 1574–1585. [Google Scholar] [CrossRef] [Green Version]
- Michaloudi, E.; Papakostas, S.; Stamou, G.; Neděla, V.; Tihlaříková, E.; Zhang, W.; Declerck, S.A.J. Reverse Taxonomy Applied to the Brachionus Calyciflorus Cryptic Species Complex: Morphometric Analysis Confirms Species Delimitations Revealed by Molecular Phylogenetic Analysis and Allows the (Re)Description of Four Species. PLoS ONE 2018, 13, e0203168. [Google Scholar] [CrossRef] [PubMed]
- Paraskevopoulou, S.; Tiedemann, R.; Weithoff, G. Differential Response to Heat Stress among Evolutionary Lineages of an Aquatic Invertebrate Species Complex. Biol. Lett. 2018, 14, 20180498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Lemmen, K.D.; Zhou, L.; Papakostas, S.; Declerck, S.A. Patterns of Differentiation in the Life History and Demography of Four Recently Described Species of the Brachionus Calyciflorus Cryptic Species Complex. Freshw. Biol. 2019, 64, 1994–2005. [Google Scholar] [CrossRef] [Green Version]
- Jeong, C.-B.; Won, E.-J.; Kang, H.-M.; Lee, M.-C.; Hwang, D.-S.; Hwang, U.-K.; Zhou, B.; Souissi, S.; Lee, S.-J.; Lee, J.-S. Microplastic Size-Dependent Toxicity, Oxidative Stress Induction, and p-JNK and p-P38 Activation in the Monogonont Rotifer (Brachionus Koreanus). Environ. Sci. Technol. 2016, 50, 8849–8857. [Google Scholar] [CrossRef]
- Rauchschwalbe, M.-T.; Fueser, H.; Traunspurger, W.; Höss, S. Bacterial Consumption by Nematodes Is Disturbed by the Presence of Polystyrene Beads: The Roles of Food Dilution and Pharyngeal Pumping. Environ. Pollut. 2021, 273, 116471. [Google Scholar] [CrossRef]
- Ramos-RodrÍguez, E.; Conde-Porcuna, J.M. Nutrient Limitation on a Planktonic Rotifer: Life History Consequences and Starvation Resistance. Limnol. Oceanogr. 2003, 48, 933–938. [Google Scholar] [CrossRef]
- Stemberger, R.S. A General Approach to the Culture of Planktonic Rotifers. Can. J. Fish. Aquat. Sci. 1981, 38, 721–724. [Google Scholar] [CrossRef]
- Bogdan, K.G.; Gilbert, J.J. Body Size and Food Size in Freshwater Zooplankton. Proc. Natl. Acad. Sci. USA 1984, 81, 6427–6431. [Google Scholar] [CrossRef] [Green Version]
- Pagano, M. Feeding of Tropical Cladocerans (Moina Micrura, Diaphanosoma Excisum) and Rotifer (Brachionus Calyciflorus) on Natural Phytoplankton: Effect of Phytoplankton Size-Structure. J. Plankton Res. 2008, 30, 401–414. [Google Scholar] [CrossRef] [Green Version]
- Coppock, R.L.; Galloway, T.S.; Cole, M.; Fileman, E.S.; Queirós, A.M.; Lindeque, P.K. Microplastics alter feeding selectivity and faecal density in the copepod, Calanus Helgolandicus. Sci. Total Environ. 2019, 687, 780–789. [Google Scholar] [CrossRef]
- Mueller, M.-T.; Fueser, H.; Trac, L.N.; Mayer, P.; Traunspurger, W.; Ho, S. Surface-Related Toxicity of Polystyrene Beads to Nematodes and the Role of Food Availability. Environ. Sci. Technol. 2020, 54, 1790–1798. [Google Scholar] [CrossRef] [PubMed]
- Rothhaupt, K.O. Algal Nutrient Limitation Affects Rotifer Growth Rate but Not Ingestion Rate. Limnol. Oceanogr. 1995, 40, 1201–1208. [Google Scholar] [CrossRef]
- Sarma, S.S.S.; Gulati, R.D.; Nandini, S. Factors Affecting Egg-Ratio in Planktonic Rotifers. Hydrobiologia 2005, 546, 361–373. [Google Scholar] [CrossRef]
- Schälicke, S.; Teubner, J.; Martin-Creuzburg, D.; Wacker, A. Fitness Response Variation within and among Consumer Species Can Be Co-Mediated by Food Quantity and Biochemical Quality. Sci. Rep. 2019, 9, 16126. [Google Scholar] [CrossRef] [PubMed]
- Osenberg, C.W.; Mittelbach, G.G. The Relative Importance of Resource Limitation and Predator Limitation in Food Chains; Polis, G.A., Winemiller, K.O., Eds.; Food Webs; Springer: Boston, MA, USA, 1996. [Google Scholar] [CrossRef]
- Devetter, M.; Sed’a, J. Decline of Clear-Water Rotifer Populations in a Reservoir: The Role of Resource Limitation. Hydrobiologia 2005, 546, 509–518. [Google Scholar] [CrossRef]
- Korez, Š.; Gutow, L.; Saborowski, R. Feeding and Digestion of the Marine Isopod Idotea Emarginata Challenged by Poor Food Quality and Microplastics. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 226, 108586. [Google Scholar] [CrossRef]
- Trotter, B.; Wilde, M.V.; Brehm, J.; Dafni, E.; Aliu, A.; Arnold, G.J.; Fröhlich, T.; Laforsch, C. Long-Term Exposure of Daphnia Magna to Polystyrene Microplastic (PS-MP) Leads to Alterations of the Proteome, Morphology and Life-History. Sci. Total Environ. 2021, 795, 148822. [Google Scholar] [CrossRef]
- Müller-Navarra, D.C.; Brett, M.T.; Liston, A.M.; Goldman, C.R. A Highly Unsaturated Fatty Acid Predicts Carbon Transfer between Primary Producers and Consumers. Nature 2000, 403, 74–77. [Google Scholar] [CrossRef]
- Ogonowski, M.; Schür, C.; Jarsén, Å.; Gorokhova, E. The Effects of Natural and Anthropogenic Microparticles on Individual Fitness in Daphnia Magna. PLoS ONE 2016, 11, e0155063. [Google Scholar] [CrossRef] [PubMed]
- Imhof, H.K.; Rusek, J.; Thiel, M.; Wolinska, J.; Laforsch, C. Do Microplastic Particles Affect Daphnia Magna at the Morphological, Life History and Molecular Level? PLoS ONE 2017, 12, e0187590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilhermino, L.; Martins, A.; Cunha, S.; Fernandes, J.O. Long-Term Adverse Effects of Microplastics on Daphnia Magna Reproduction and Population Growth Rate at Increased Water Temperature and Light Intensity: Combined Effects of Stressors and Interactions. Sci. Total Environ. 2021, 784, 147082. [Google Scholar] [CrossRef] [PubMed]
- Schälicke, S.; Sobisch, L.; Martin-Creuzburg, D.; Wacker, A. Food Quantity–Quality Co-limitation: Interactive Effects of Dietary Carbon and Essential Lipid Supply on Population Growth of a Freshwater Rotifer. Freshw Biol 2019, 64, 903–912. [Google Scholar] [CrossRef]
- Rothhaupt, K. Differences in Particle Size-Dependent Feeding Efficiencies of Closely Related Rotifer Species. Limnol. Oceanogr. 1990, 35, 16–23. [Google Scholar] [CrossRef]
- Drago, C.; Pawlak, J.; Weithoff, G. Biogenic Aggregation of Small Microplastics Alters Their Ingestion by a Common Freshwater Micro-Invertebrate. Front. Environ. Sci. 2020, 8, 264. [Google Scholar] [CrossRef]
- Starkweather, P.L. Aspects of the Feeding Behavior and Trophic Ecology of Suspension-Feeding Rotifers. Hydrobiologia 2004, 73, 63–72. [Google Scholar] [CrossRef]
- Starkweather, P.L.; Gilbert, J.J.; Frost, T.M. Bacterial Feeding by the Rotifer Brachionus calyciflorus: Clearance and Ingestion Rates, Behavior and Population Dynamics. Oecologia 1979, 44, 26–30. [Google Scholar] [CrossRef]
- Kirk, K.L.; Gilbert, J.J. Suspended Clay and the Population Dynamics of Planktonic Rotifers and Cladocerans. Ecology 1990, 71, 1741–1755. [Google Scholar] [CrossRef]
- Kirk, K.L. Inorganic Particles Alter Competition in Grazing Plankton: The Role of Selective Feeding. Ecology 1991, 72, 915–923. [Google Scholar] [CrossRef]
- Schür, C.; Zipp, S.; Thalau, T.; Wagner, M. Microplastics but Not Natural Particles Induce Multigenerational Effects in Daphnia Magna. Environ. Pollut. 2020, 260, 113904. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.-P.; Cole, M.; Chan, B.K.K. Review: Effects of microplastic on zooplankton survival and sublethal responses. In Oceanography and Marine Biology: An Annual Review; Hawkins, S.J., Allcock, A.L., Bates, A.E., Evans, A.J., Firth, L.B., McQuaid, C.D., Russell, B.D., Smith, I.P., Swearer, S.E., Todd, P.A., Eds.; Editors Taylor and Francis; CRC Press: London, UK, 2020; Volume 58, pp. 351–393. ISBN 978-0-429-35149-5. [Google Scholar]
- Klein, K.; Heß, S.; Nungeß, S.; Schulte-Oehlmann, U.; Oehlmann, J. Particle Shape Does Not Affect Ingestion and Egestion of Microplastics by the Freshwater Shrimp Neocaridina Palmata. Env. Sci Pollut Res. 2021, 28, 62246–62254. [Google Scholar] [CrossRef]
- Botterell, Z.L.R.; Beaumont, N.; Cole, M.; Hopkins, F.E.; Steinke, M.; Thompson, R.C.; Lindeque, P.K. Bioavailability of Microplastics to Marine Zooplankton: Effect of Shape and Infochemicals. Environ. Sci. Technol. 2020, 54, 12024–12033. [Google Scholar] [CrossRef] [PubMed]
- Desforges, J.-P.W.; Galbraith, M.; Ross, P.S. Ingestion of Microplastics by Zooplankton in the Northeast Pacific Ocean. Arch Env. Contam Toxicol 2015, 69, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Besseling, E.; Redondo-Hasselerharm, P.; Foekema, E.M.; Koelmans, A.A. Quantifying Ecological Risks of Aquatic Micro- and Nanoplastic. Crit. Rev. Environ. Sci. Technol. 2019, 49, 32–80. [Google Scholar] [CrossRef] [Green Version]
- Lindeque, P.K.; Cole, M.; Coppock, R.L.; Lewis, C.N.; Miller, R.Z.; Watts, A.J.; Galloway, T.S. Are we underestimating microplastic abundance in the marine environment? A comparison of microplastic capture with nets of different mesh-size. Environ. Pollut. 2020, 265, 114721. [Google Scholar] [CrossRef]
- Scherer, C.; Weber, A.; Stock, F.; Vurusic, S.; Egerci, H.; Kochleus, C.; Arendt, N.; Foeldi, C.; Dierkes, G.; Wagner, M.; et al. Comparative assessment of microplastics in water and sediment of a large European river. Sci. Total. Environ. 2020, 738, 139866. [Google Scholar] [CrossRef]
- Weithoff, G.; Lorke, A.; Walz, N. Effects of Water-Column Mixing on Bacteria, Phytoplankton, and Rotifers under Different Levels of Herbivory in a Shallow Eutrophic Lake. Oecologia 2000, 125, 91–100. [Google Scholar] [CrossRef]
- Zimmermann, L.; Göttlich, S.; Oehlmann, J.; Wagner, M.; Völker, C. What Are the Drivers of Microplastic Toxicity? Comparing the Toxicity of Plastic Chemicals and Particles to Daphnia Magna. Environ. Pollut. 2020, 267, 115392. [Google Scholar] [CrossRef]
- Zimmermann, L.; Dierkes, G.; Ternes, T.A.; Völker, C.; Wagner, M. Benchmarking the in Vitro Toxicity and Chemical Composition of Plastic Consumer Products. Environ. Sci. Technol. 2019, 53, 11467–11477. [Google Scholar] [CrossRef] [Green Version]
- Muncke, J. Exposure to Endocrine Disrupting Compounds via the Food Chain: Is Packaging a Relevant Source? Sci. Total Environ. 2009, 407, 4549–4559. [Google Scholar] [CrossRef] [PubMed]
- Lithner, D.; Nordensvan, I.; Dave, G. Comparative Acute Toxicity of Leachates from Plastic Products Made of Polypropylene, Polyethylene, PVC, Acrylonitrile–Butadiene–Styrene, and Epoxy to Daphnia Magna. Environ. Sci. Pollut. Res. 2012, 19, 1763–1772. [Google Scholar] [CrossRef]
- Schrank, I.; Trotter, B.; Dummert, J.; Scholz-Böttcher, B.M.; Löder, M.G.J.; Laforsch, C. Effects of Microplastic Particles and Leaching Additive on the Life History and Morphology of Daphnia magna. Environ. Pollut. 2019, 255, 113233. [Google Scholar] [CrossRef]
- Weisse, T.; Laufenstein, N.; Weithoff, G. Multiple Environmental Stressors Confine the Ecological Niche of the Rotifer Cephalodella Acidophila. Freshw. Biol. 2013, 58, 1008–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiltunen, M.; Vehniäinen, E.-R.; Kukkonen, J.V.K. Interacting Effects of Simulated Eutrophication, Temperature Increase, and Microplastic Exposure on Daphnia. Environ. Res. 2021, 192, 110304. [Google Scholar] [CrossRef]
- Aljaibachi, R.; Callaghan, A. Impact of Polystyrene Microplastics on Daphnia magna Mortality and Reproduction in Relation to Food Availability. PeerJ 2018, 6, e4601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aljaibachi, R.; Laird, W.B.; Stevens, F.; Callaghan, A. Impacts of Polystyrene Microplastics on Daphnia magna: A Laboratory and a Mesocosm Study. Sci. Total Environ. 2020, 705, 135800. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Egg-Ratio | Probability of Survival | |||||
|---|---|---|---|---|---|---|
| Independent variables | Df | F-Value | p-Value | Df | F-Value | p-Value |
| Alg | 1137 | 125.5 | <0.0001 | 1137 | 0.4 | 0.534 |
| food | 1137 | 997.0 | <0.0001 | 1137 | 28.6 | <0.0001 |
| food × Alg | 1137 | 33.5 | <0.0001 | 1137 | 2.8 | 0.099 |
| food × Treatment | 5137 | 1.0 | 0.422 | 5137 | 3.9 | <0.01 |
| Specie | 1137 | 1.3 | 0.258 | 1137 | 20.2 | <0.0001 |
| Specie × food | 1137 | 16.6 | <0.0001 | 1137 | 2.4 | 0.126 |
| Specie × food × Treatment | 5137 | 1.5 | 0.190 | 5137 | 0.6 | 0.699 |
| Specie × Treatment | 5137 | 0.3 | 0.907 | 5137 | 3.3 | <0.01 |
| Treatment | 5137 | 20.3 | <0.0001 | 5137 | 5.6 | <0.001 |
| Treatment × Alg | 5137 | 4.2 | <0.01 | 5137 | 3.2 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drago, C.; Weithoff, G. Variable Fitness Response of Two Rotifer Species Exposed to Microplastics Particles: The Role of Food Quantity and Quality. Toxics 2021, 9, 305. https://doi.org/10.3390/toxics9110305
Drago C, Weithoff G. Variable Fitness Response of Two Rotifer Species Exposed to Microplastics Particles: The Role of Food Quantity and Quality. Toxics. 2021; 9(11):305. https://doi.org/10.3390/toxics9110305
Chicago/Turabian StyleDrago, Claudia, and Guntram Weithoff. 2021. "Variable Fitness Response of Two Rotifer Species Exposed to Microplastics Particles: The Role of Food Quantity and Quality" Toxics 9, no. 11: 305. https://doi.org/10.3390/toxics9110305
APA StyleDrago, C., & Weithoff, G. (2021). Variable Fitness Response of Two Rotifer Species Exposed to Microplastics Particles: The Role of Food Quantity and Quality. Toxics, 9(11), 305. https://doi.org/10.3390/toxics9110305