Impacts of Neonicotinoids on Molluscs: What We Know and What We Need to Know


Abstract
:1. Introduction
2. Neonicotinoids
2.1. Chemical Properties, Registration, Use and Efficacy

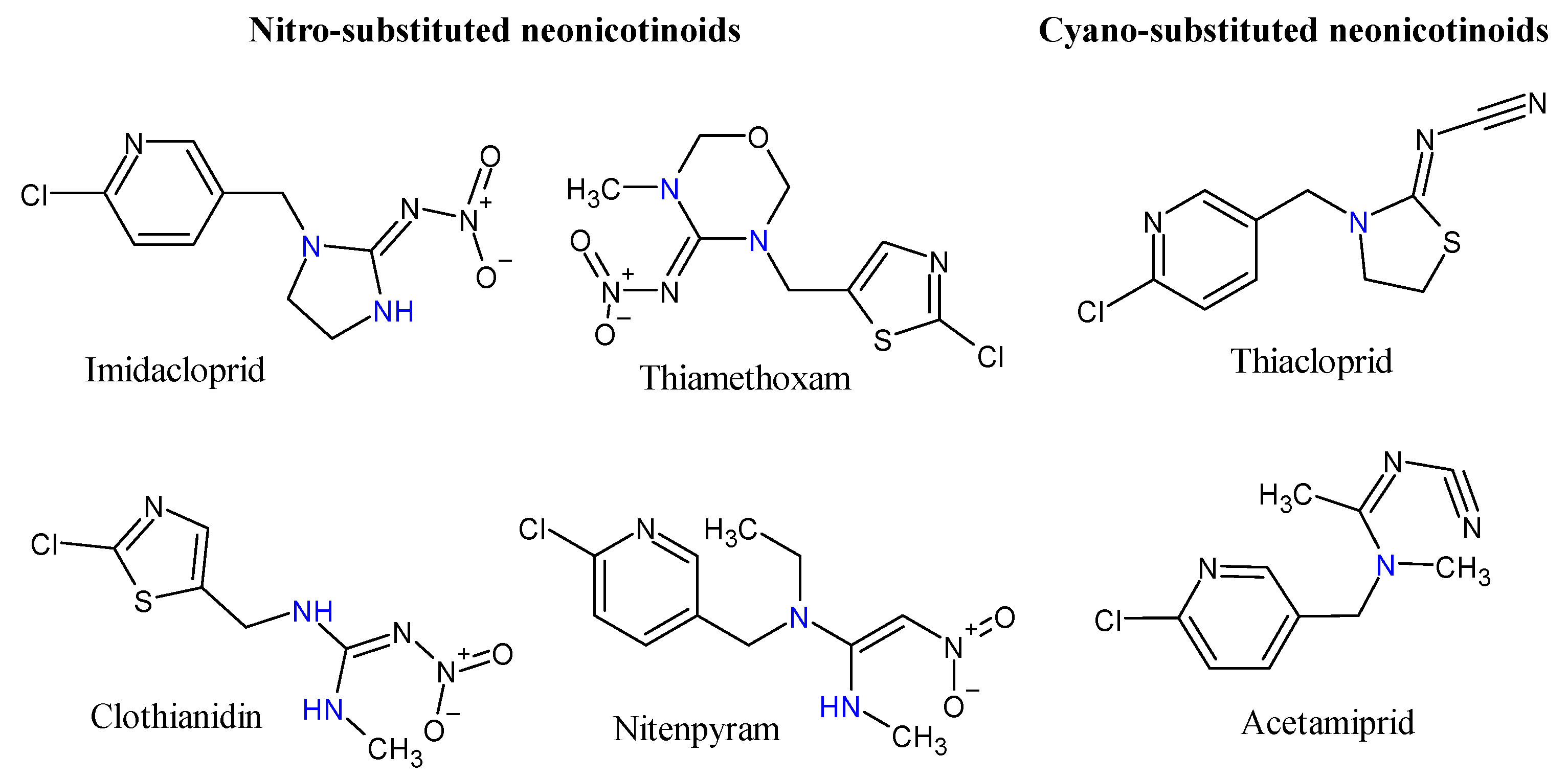{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neonicotinoids Properties | Imidacloprid | Acetamiprid | Nitenpyram | Thiamethoxam | Thiacloprid | Clothianidin |
|---|---|---|---|---|---|---|
| Released year | 1991 | 1995 | 1995 | 1998 | 2000 | 2002 |
| Molecular formula | C9H10ClN5O2 | C10H11ClN4 | C11H15ClN4O2 | C8H10ClN5O3S | C10H9ClN4S | C6H8ClN5O2S |
| Molecular weight (g/mol) | 255.7 | 222.67 | 270.72 | 291.71 | 252.72 | 249.68 |
| Vapour Pressure (mm Hg) | 1 × 10−07 | 4.5 × 10−05 | 8.2 × 10−12 | 6.6 × 10−06 | 6 × 10−12 | 1.3 × 10−07 |
| Hydrolysis half-life at pH 7 (days) | >2000 | na | na | ≥572 | 10 to 63 | na |
| Octanol-water coefficient (Kow) | 3.7 | 6.27 | −0.66 | −0.13 | 1.26 | 5 |
| Henry’s constant (atm m3/mole) | 6.5 × 10−11 | 7.92 × 10−08 | 7.9 × 10−11 | 4.7 × 10−10 | 1.08 × 10−14 | 2.9 × 10−16 |
| Melting point (°C) | 136.4 to 143.8 | 98.9 | ~82.8 | 139.1 | 136 | 176.8 |
| Anaerobic aquatic half-life (days) | 27.1 | 45 | ~3 | 35.5 | >365 | 27 |
| Aqueous photolysis half-life (hours) | 1 to 4 | >34 | ~4.4 | ≥3.36 | 42 | <24 |
| Water solubility (mg/L at 20 °C) | 510 to 610 | 4200 | 5.7 × 10+05 | 4100 | 185 | 327 |
| Soil photolysis half-life (days) | 38.9 | 25.1 | 1 to 15 | 47 to 54 | na | 34 |
| Field dissipation half-life (days) | 26.5–229 | <18 | <4 | 72 to 111 | 19 | 2 to 27 |
| Soil adsorption coefficient (Kd) | 0.956–4.18 | <4.1 | na | 0.59 to 2.03 | na | 0.62 to 1.94 |
| Trade names | Confidor Merit Gaucho Admire Kohinor Prothor Advantage Gaucho Spectrum Premise Winner | Assail Intruder Adjust Rescate Tristar Saurus Prize Tristar Mosiplan Gazelle Trivor | Capstar Bestguard | Actara Cruiser Helix Platinum Centric Maxide Meridian Flashship Endigo Optigard Durivo Agri-flex Voliam | Calypso Biscaya Viper Piranha | Aloft Clutch Arena Votico Prosper Belay |
2.2. Environmental Fate
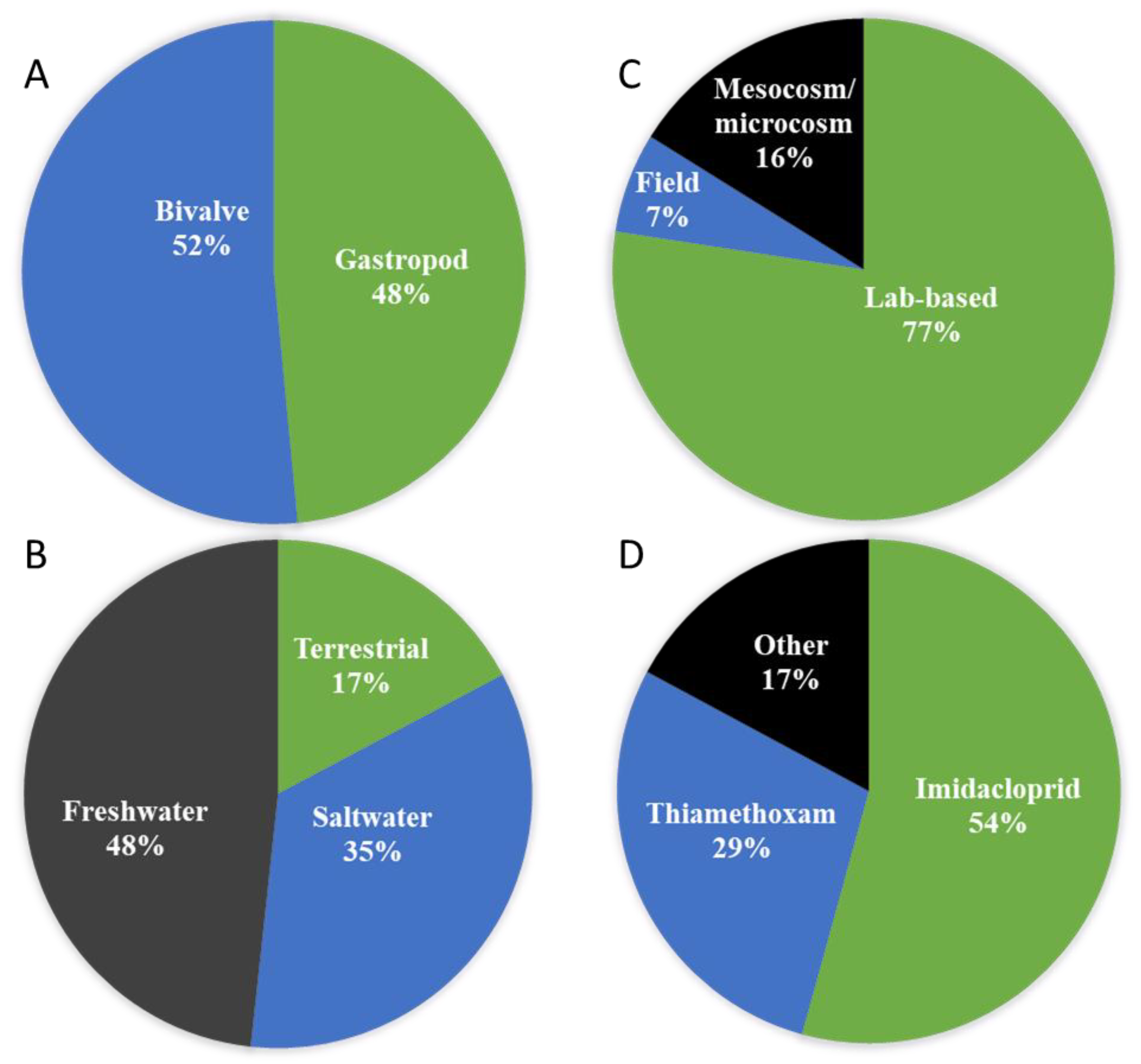
3. A Literature Search of Neonicotinoids Studies on Molluscs
4. Impacts of Neonicotinoids in Molluscs: What Do We Know
| Species (Life Stage) | Species Class (Habitat) | Location (Study Type) | Neonicotinoids | Dose (Duration of Exposure) | Findings | Reference |
|---|---|---|---|---|---|---|
| Accumulation | ||||||
| Saccostrea glomerata (adult) | Bivalve (saltwater) | Australia (lab-based) | Imidacloprid and a formulation (spectrum 200SC) | 0.01, 0.05, 0.1, 1 and 2 mg/L (2 weeks) | Accumulated in the gill (0.40 µg/g at 0.01 mg/L and 4.6 µg/g at 2 mg/L exposures), adductor muscle (0.41 µg/g at 0.01 mg/L and 7.14 µg/g at 2 mg/L exposures) and digestive gland (0.22 µg/g at 0.01 mg/L and 4.39 µg/g at 2 mg/L exposures) | Ewere et al. [31] |
| Saccostrea glomerata (adult) | Bivalve (saltwater) | Australia (lab-based) | Imidacloprid | 0.2 mg/L (1 to 3 days) | Accumulated in the gill (0.7 µg/g), adductor muscle (1.2 µg/g) and digestive gland (0.4 µg/g). | Ewere et al. [30] |
| Deroceras reticulatum (*) | Gastropod (terrestrial) | USA (lab- and field-based) | Thiamethoxam and a formulation (CruiserMaxx®) | 0.08 and 0.15 mg/seed (8 days) | Accumulated residues and up to 0.5 µg/g in field-collected samples. | Douglas et al. [29] |
| Behaviour | ||||||
| Helix aspersa (adult) | Gastropod (terrestrial) | Algeria (lab-based) | Thiamethoxam | 100 and 200 mg/L (1 week) | Significant inhibition of locomotion and feeding at either concentration | Hamlet et al. [91] |
| Crassostrea gigas (larvae) | Bivalve (saltwater) | France (lab-based) | Imidacloprid | 20 µg/L (24 h) | No effect on locomotion | Kuchovská et al. [92] |
| Deroceras reticulatum, Arion distinctus and Milax gagates (adults) | Gastropod (terrestrial) | UK (lab- and field-based) | Imidacloprid formulation (Gaucho) | 0.7, 1.4, 2.8 g/kg seed (4 to 11 days) | Significant reduction in feeding on winter wheat at ≥2.8 g/kg (lab-based) and ≥0.7 g/kg (field-based) | Simms et al. [93] |
| Corbicular fluminea (larvae) | Bivalve (freshwater) | China (lab-based) | Imidacloprid | 0.02, 0.2 and 2 mg/L (30 days) | Inhibition of feeding at 2 mg/L and burrowing at 0.02 mg/L exposures | Shan et al. [89] |
| Unio tumidus (adult) | Bivalve (saltwater) | Poland (lab-based) | Thiacloprid | 10 µg/L (168 h) | Reduction of shell opening level and increase of shell opening rate | Chmist et al. [94] |
| Saccostrea glomerata (adult) | Bivalve (saltwater) | Australia (lab-based) | Imidacloprid | 0.125 to 2 mg/L (1 to 4 days) | Reduction of filtration rate at 2 mg/L (day 1) and 0.5 and 1 mg/L (day 4) | Ewere et al. [30] |
| Physiology | ||||||
| Immunity | ||||||
| Mytilus galloprovincialis (adult) | Bivalve (saltwater) | Italy (lab-based) | Imidacloprid formulation (Calypso 480 SC) | 7.77 and 77.7 mg/L (96 h and 20 days) | Increased hemocytes mortality rate, and reduction in hemolymph Cl− and Na+ | Stara et al. [32] |
| Crassostrea gigas (larvae) | Bivalve (saltwater) | Australia (lab-based) | Imidacloprid | 0.001 mg/L (53 h) | No increased susceptibility to disease caused by OsHV-1 | Oliver et al. [95] |
| Saccostrea glomerata (adult) | Bivalve (saltwater) | Australia (lab-based) | Imidacloprid | 0.01 to 1 mg/L (96 h) | Increase in hemocyte counts and decrease in hemocytes aggregation at ≥0.1 mg/L exposure, but no effect of phagocytosis and hemocytes different types | Ewere et al. [35] |
| Growth and morphology | ||||||
| Marisa cornuarietis (embryo) | Gastropod (freshwater) | Germany (lab-based) | Imidacloprid | Up to 50 mg/L (9 days | No effect on the formation of eye and tentacles, hatching, as well as weight post-hatching | Sawasdee and Köhler [96] |
| Crassostrea gigas (larvae) | Bivalve (saltwater) | France (lab-based) | Imidacloprid | 0.2–2000 µg/L (30 h) | Increased in percentage of abnormal larvae at ≥200 µg/L | Kuchovská et al. [92] |
| Saccostrea glomerata (adult) | Bivalve (saltwater) | Australia (lab-based) | Imidacloprid and a formulation (spectrum 200SC) | ≤2 mg/L (2 weeks) | No significant effect on the condition index | Ewere et al. [31] |
| Crassostrea virginica (*) | Bivalve (saltwater) | Canada (flow-through) | Thiamethoxam | 119 mg/L (96 h) | No significant inhibition of shell growth | Finnegan et al. [97] |
| Planorbella pilsbryi (juvenile) and Lampsilis fasciola (juvenile) | Gastropod and bivalve (freshwater) | Canada (lab-based) | Imidacloprid and thiamethoxam | 0.001 to 1 mg/L (28 days) | Significant reduction of growth and biomass production at ≥21 µg/L (imidacloprid) and ≥24.8 µg/L (thiamethoxam) | Prosser et al. [98] |
| Histopathology | ||||||
| Helix aspersa (adult) | Gastropod (terrestrial) | Algeria (lab-based) | Thiamethoxam | 10, 20 and 40 mg/L (20 days) | Degeneration of digestive tubules and breakdown of basement membrane in the hepatopancreas in all concentrations tested | Hamlet et al. [99] |
| Parreysia cylindrica | Bivalve (freshwater) | Indian (lab-based) | Thiamethoxam | 14 mg/L (24 h, 96 h and 7 days) and 2.8 mg/L (21 days) | hypertrophy and sloughing of the epithelium, epithelial necrosis, tubular hyperplasia and rupture of the epithelial layer ≥14 mg/L. After 21 days, epithelial cells separated from the basement membrane | Patil [100] |
| Mytilus galloprovincialis (adult) | Bivalve (saltwater) | Italy (lab-based) | Imidacloprid formulation (Calypso 480 SC) | 7.77 and 77.7 mg/L (10 and 20 days) | Caused alteration in the gills and digestive gland at the concentrations tested | Stara et al. [32] |
| Corbicular fluminea (larvae) | Bivalve (freshwater) | China (lab-based) | Imidacloprid | 0.02, 0.2 and 2 mg/L (30 days) | Gill and digestive tissue damage at ≥0.02 mg/L, with severe damage at ≥2 mg/L | Shan et al. [89] |
| Helix aspersa (adult) | Gastropod (terrestrial) | Algeria (lab-based) | Thiamethoxam | 25–200 mg/L (6 weeks) | Increase in the number of excretory vacuoles, breakdown of basement membrane and degeneration of digestive cells of the hepatopancreas at ≥25 mg/L | Hamlet et al. [91] |
| Biochemistry | ||||||
| Synaptic connection | ||||||
| Lymnaea stagnalis (larvae) | Gastropod (freshwater) | The Netherlands (lab-based) | Imidacloprid | 0.001, 0.01 and 0.1 mg/L (10 days) | Increase in choline and acetylcholine turnover at ≥0.01 mg/L | Tufi et al. [101] |
| Lymnaea stagnalis | Gastropod (freshwater) | Japan (lab-based | Imidacloprid | na | Significant low affinity to the nicotinic acetylcholine receptor, possibly due to the presence of two orientation on the receptor for imidacloprid binding | Tomizawa and Casida [102] |
| Lymnaea stagnalis | Gastropod (freshwater) | Hungary (lab-based) | Acetamiprid formulations (Mospilan), imidacloprid formulation (Kohinor), thiamethoxam (Actara) and thiacloprid (Calypso) | 0.01 and 0.1 mg/mL (5 s) | Each of the neonicotinoids inhibited the VD4-RPeD1. Calypso block 90% of excitatory postsynaptic potentials | Vehovszky et al. [103] |
| Cholinesterase | ||||||
| Mytilus galloprovincialis (adult) | Bivalve (saltwater) | Italy (lab-based) | Imidacloprid and thiacloprid | 0.1, 1 and 10 mg/L (96 h) | Reduction of acetylcholinesterase activity in the gill at ≥0.1 mg/L imidacloprid exposure and thiacloprid caused induction of acetylcholinesterase activity at 1 mg/L exposure and inhibition at 0.1 and 10 mg/L exposure | Dondero et al. [104] |
| Helix aspersa (adult) | Gastropod (terrestrial) | Algeria (lab-based) | Thiamethoxam | 25–200 mg/L (6 weeks) | Inhibition of acetylcholinesterase activity at ≥25 mg/L, with higher concentrations causing greater inhibition | Smina et al. [90] |
| Saccostrea glomerata (adult) | Bivalve (saltwater) | Australia (lab-based) | Imidacloprid and formulation | 0.01, 0.05, 0.1, 1 and 2mg/L (2 weeks) | Reduction of acetylcholinesterase in the gill at 2 mg/L | Ewere et al. [31] |
| Biomphalaria straminea (adult) | Gastropod (freshwater) | Argentina (lab-based) | Acetamiprid and a formulation (Assail 70® WP) | 150 and 1500 µg/L (14 days) | No effect on cholinesterase activity | Cossi et al. [105] |
| Saccostrea sp. (adult) | Bivalve (saltwater) | Colombia (lab-based) | Imidacloprid formulation (Imidogen 350 SC) | 0.1, 1, 10 and 100 mg/L (96 h) | Reduction in total cholinesterase and eserine-sensitive cholinesterase activity in gill at ≥10 mg/L, and increase in eserine-sensitive cholinesterase activity in the digestive gland at 100 mg/L. Reduction of eserine-resistance cholinesterase activity in the adductor muscle at 10 mg/L | Moncaleano-Niño et al. [106] |
| Corbicular fluminea (larvae) | Bivalve (freshwater) | China (lab-based) | Imidacloprid | 0.2, 0.2 and 2 mg/L (30 days) | Reduction of acetylcholinesterase in the gill 2 mg/L and digestive gland at ≥0.2 mg/L | Shan et al. [89] |
| Antioxidant activity | ||||||
| Corbicular fluminea (larvae) | Bivalve (freshwater) | China (lab-based) | Imidacloprid | 0.02, 0.2 and 2 mg/L (30 days) | Increase in glutathione S-transferase, catalase and superoxide dismutase activities in the gill and digestive gland at ≥0.02 mg/L | Shan et al. [89] |
| Saccostrea glomerata (adult) | Bivalve (saltwater) | Australia (lab-based) | Imidacloprid | 0.01, 0.1 and 1 mg/L (96 h) | Increase in hemolymph glutathione S-transferase activity at ≥0.1 mg/L | Ewere et al. [35] |
| Helix aspersa (adult) | Gastropod (terrestrial) | Algeria (lab-based) | Thiamethoxam | 25–200 mg/L (6 weeks) | Induction of glutathione S-transferase and catalase activity at ≥25 mg/L, with higher concentrations causing greater induction | Smina et al. [90] |
| Biomphalaria straminea (adult) | Gastropod (freshwater) | Argentina (lab-based) | Acetamiprid and a formulation (Assail 70® WP) | 150 and 1500 µg/L (14 days) | Significant increase in carboxylase activity, glutathione S-transferase activity, glutathione content, and decrease/inhibition of catalase activity, oxygen species levels and superoxide dismutase at both concentrations | Cossi et al. [105] |
| Mytilus galloprovincialis (adult) | Bivalve (saltwater) | Italy (lab-based) | Imidacloprid formulation (Calypso 480 SC) | 7.77 and 77.7 mg/L (20 days) | Significant reduction of superoxide dismutase in the digestive gland and reduction of catalase activity in the gills at both concentrations | Stara et al. [32] |
| Saccostrea glomerata (adult) | Bivalve (saltwater) | Australia (lab-based) | Imidacloprid and formulation (spectrum 200SC) | 0.01, 0.05, 0.1, 1 and 2mg/L (2 weeks) | Reduction of catalase in the digestive gland at 0.1 mg/L and glutathione S-transferase activities in the gill and digestive gland at ≥0.01 mg/L | Ewere et al. [31] |
| Energy reserve | ||||||
| Helix aspersa (*) | Gastropod (terrestrial) | Algeria (lab-based) | Thiamethoxam | 25–200 mg/L (6 weeks) | Reduction of total lipid content at ≥100 mg/L | Hamlet et al. [91] |
| Saccostrea glomerata (adult) | Bivalve (saltwater) | Australia (lab-based) | Imidacloprid and formulation (spectrum 200SC) | 0.01, 0.05, 0.1, 1 and 2mg/L (2 weeks) | Altered the ratios and major classes of fatty acids at ≥0.01 mg/L | Ewere et al. [31] |
| Lymnaea stagnalis (larvae) | Gastropod (freshwater) | The Netherlands (lab-based) | Imidacloprid | 0.001, 0.01 and 0.1 mg/L (10 days) | Decrease of fatty acids, possibly due to a downregulation of fatty acids biosynthesis. Exposure also caused an upregulation of lipids at ≥0.01 mg/L | Tufi et al. [101] |
| Helix aspersa (*) | Gastropod (terrestrial) | Algeria (lab-based) | Thiamethoxam | 25–200 mg/L (6 weeks) | Reduction of tissue carbohydrate and protein contents at ≥100 mg/L | Hamlet et al. [91] |
| Omics | ||||||
| Mytilus galloprovincialis (adult) | Bivalve (saltwater) | Italy (lab-based) | Imidacloprid and thiacloprid | 2 mg/L (96 h) | Upregulation of heat shock proteins gene, protein translation genes, and downregulation of chitinase, endo-beta-glucanase, scavenger receptor cysteine-rich partial and profoldin subunit 4 | Dondero et al. [104] |
| Corbicular fluminea (larvae) | Bivalve (freshwater) | China (lab-based) | Imidacloprid | 0.02, 0.2 and 2 mg/L (30 days) | Downregulation of multixenobiotic resistance and heat shock protein genes at ≥0.02 mg/L | Shan et al. [89] |
| Saccostrea glomerata (adult) | Bivalve (saltwater) | Australia (lab-based) | Imidacloprid | 2mg/L (96 h) | Upregulation of cargo and scavenger receptor activity-related genes and downregulation genes involved in axoneme, cilium or flagellum-dependent cell motility, dephosphorylation and phosphatase activity | Ewere et al. [30] |
| Crassostrea gigas (larvae) | Bivalve (saltwater) | France (lab-based) | Imidacloprid | 0.2–2000 µg/L (72 h) | Upregulation of SOD [Cu/Zn], genes coding for two metallothioneins (mt1 and mt2), and downregulation of SOD [Mn], genes linked with apoptosis and cell cycle regulation at ≥10 µg/L | Kuchovská et al. [92] |
| Saccostrea glomerata (adult) | Bivalve (saltwater) | Australia (lab-based) | Imidacloprid | 0.01, 0.1 and 1mg/L (96 h) | Expression of several hemolymph proteins, including the upregulation of severin, heat shock proteins, superoxide dismutase and calmodulin, and the downregulation of collagens, actins, myosin heavy chain and CEP209_CC5 domain-containing protein ≥0.01 mg/L | Ewere et al. [35] |
| Population dynamics | ||||||
| Marisa cornuarietis (embryo) | Gastropod (freshwater) | Germany (lab-based) | Imidacloprid | 50 mg/L (9 days) | No effect on mortality | Sawasdee and Köhler [96] |
| Melanoides tuberculatus (adult), Melanoides tuberculatus (juvenile), Lamellidens marginalis* and Viviparous bengalensis* | Bivalve and gastropod (freshwater) | Bangladesh (microcosm) | Imidacloprid | 0.003–3 µg/L (2–23 days) | No effect on mortality/population | Sumon et al. [107] |
| Lampsilis fasciola and Planorbella pilsbryi (juvenile) | Bivalve and gastropod (freshwater) | Canada (lab-based) | Imidacloprid, thiamethoxam, clothianidin, acetamiprid and thiacloprid | 0.01–10 mg/L (7 and 28 days) | No reduction in viability at low concentrations. The estimated 7 days LC50 for the first three neonicotinoids on the list was ≥4 mg/L, and the 28 days LC50 was ~182 µg/L | Prosser et al. [98] |
| Physella acuta* and Sphaeriidae* | Bivalve and gastropod (freshwater) | Spain (mesocosm) | Imidacloprid and neonicotinoids mixtures (containing imidacloprid, acetamiprid, thiacloprid, clothianidin and thiamethoxam) | 0.2–250 µg/L (0–56 days) | Significant increase in the number of Sphaeriidae 250 µg/L imidacloprid treatment and a decrease of Physella acuta 250 µg/L neonicotinoids mixtures | Rico et al. [108] |
| Physa sp.*, Lymnaea sp.*, Planorbis sp.* and Musculium lacustre* | Bivalve and gastropod (freshwater) | UK (mesocosm) | Thiamethoxam formulation (Actara® 25 WG) | 1–100 µg/L (0–92 days) | No effect on mortality/abundance | Finnegan et al. [109] |
| Lampsilis siliquoidea (juvenile and adult) and Villosa iris (glochidia) | Bivalve (freshwater) | Canada (lab-based) | Imidacloprid, clothianidin and thiamethoxam | 0–21 mg/L (24 h for glochidia and 28 days for juvenile and adult)) | Only 8% decrease in glochidia viability at the maximum concentration tested. Clothianidin exposure at >9 mg/L caused 22% mortality in juvenile Lampsilis siliquoidea | Salerno et al. [110] |
| Biomphalaria straminea (adult and juvenile) | Gastropod (freshwater) | Argentina (lab-based) | Acetamiprid and a formulation (Assail 70® WP) | 150 and 1500 µg/L (14 days for adult and 30 days for juvenile) | No effect on mortality | Cossi et al. [105] |
| Lymnaea stagnalis and Radix peregra (*) | Gastropod (freshwater) | Canada (lab-based) | Thiamethoxam | 100 mg/L (48 h) | No effects on mortality and immobilisation | Finnegan et al. [97] |
| Planorbella trivolvis and Physella acuta (*) | Gastropod (freshwater) | USA (mesocosm) | Clothianidin formulation (Arena) | 0.6, 5 and 352 µg/L (48 h) | No significant effect | Miles et al. [111] |
| Deroceras reticulatum (*) | Gastropod (terrestrial) | USA (field-based) | Thiamethoxam and a formulation (CruiserMaxx®) | ≥0.152 mg/seed (2 weeks after seed emerge) | Increase in population density due to reduction in predation or predators density | Douglas et al. [29] |
| Radix sp. (*) | Gastropod (freshwater) | Germany (microcosm) | Imidacloprid | 0.6–40 µg/L (7 weeks) | Increase in the population at 40 µg/L, probably due to a decrease in competition from other sensitive species | Colombo et al. [112] |
4.1. Neonicotinoids Accumulation
4.2. Impact on Behaviour
4.3. Impacts on Physiology
4.4. Impact on Biochemistry
4.5. Impacts on Population Dynamics
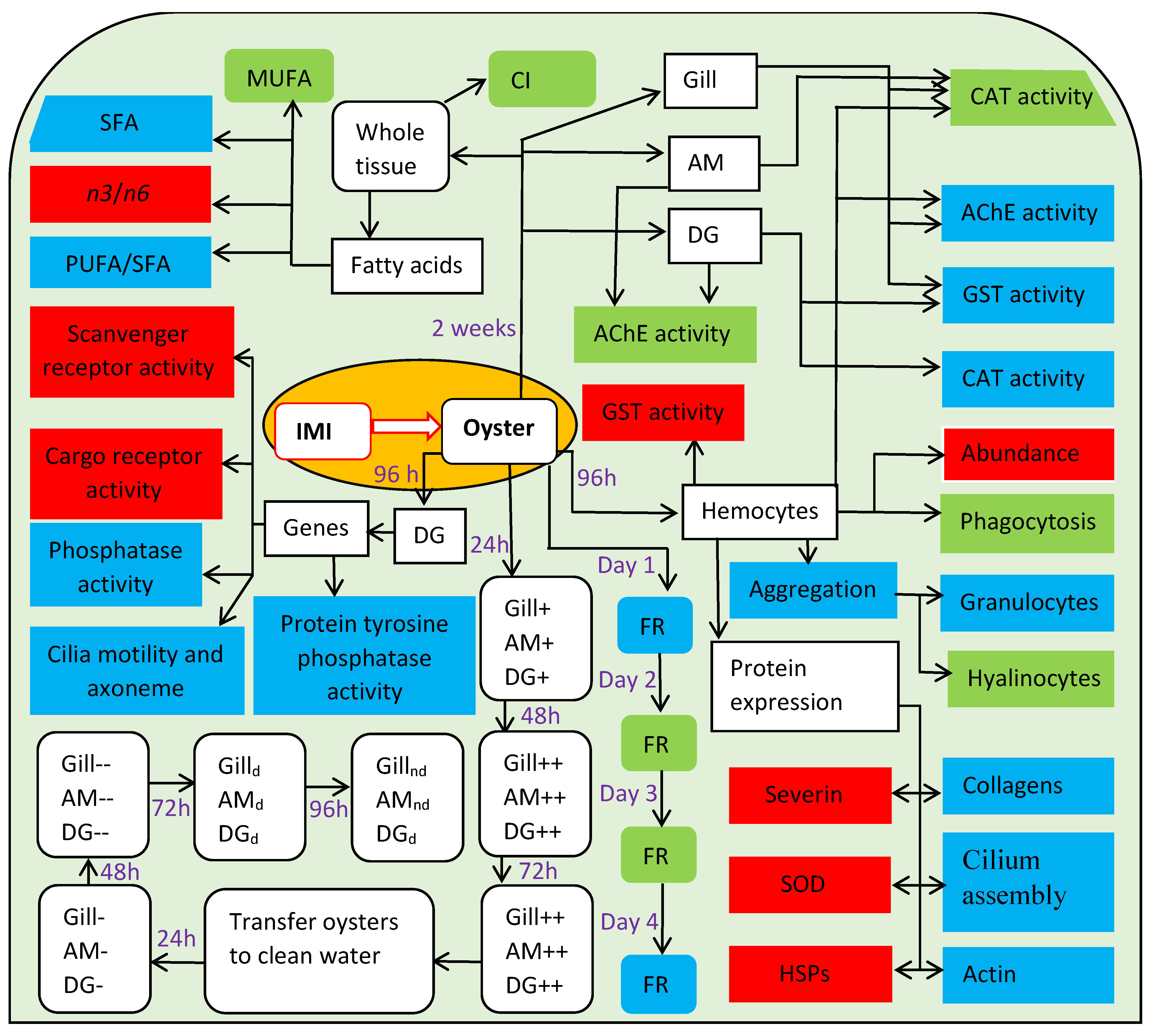
5. Synthesis and What We Still Need to Know
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Carvalho, F.P. Pesticides, environment, and food safety. Food Energy Secur. 2017, 6, 48–60. [Google Scholar]
- Mcconville, A. Why the world needs AgBioscience. Australas. Biotechnol. 2015, 25, 43. [Google Scholar]
- Popp, J.; Pető, K.; Nagy, J. Pesticide productivity and food security. A review. Agron. Sustain. Dev. 2013, 33, 243–255. [Google Scholar]
- Chagnon, M.; Kreutzweiser, D.; Mitchell, E.A.; Morrissey, C.A.; Noome, D.A.; Van Der Sluijs, J.P. Risks of large-scale use of systemic insecticides to ecosystem functioning and services. Environ. Sci. Pollut. Res. 2015, 22, 119–134. [Google Scholar]
- Stanley, J.; Preetha, G. Pesticide Toxicity to Non-Target Organisms: Exposure, Toxicity and Risk Assessment Methodologies; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Bragança, I.; Grosso, C.; Rede, D.; Sousa, S.R.; Lemos, P.C.; Domingues, V.F.; Delerue-Matos, C. Ecotoxicological Effects of Insecticides in Plants Assessed by Germination and Other Phytotoxicity Tools. In Biotic and Abiotic Stress Tolerance in Plants; Springer: Cham, Switzerland, 2018; pp. 47–76. [Google Scholar]
- O’Shea, T.; Everette, A.; Ellison, L. Cyclodiene insecticide, DDE, DDT, arsenic, and mercury contamination of big brown bats (Eptesicus fuscus) foraging at a Colorado Superfund site. Arch. Environ. Contam. Toxicol. 2001, 40, 112–120. [Google Scholar] [PubMed]
- Kumari, B.; Madan, V.; Kathpal, T. Status of insecticide contamination of soil and water in Haryana, India. Environ. Monit. Assess. 2008, 136, 239–244. [Google Scholar] [PubMed]
- Zidar, P.; Hribar, M.; Žižek, S.; Štrus, J. Behavioural response of terrestrial isopods (Crustacea: Isopoda) to pyrethrins in soil or food. Eur. J. Soil Biol. 2012, 51, 51–55. [Google Scholar]
- Li, X.; Chen, J.; He, X.; Wang, Z.; Wu, D.; Zheng, X.; Zheng, L.; Wang, B. Simultaneous determination of neonicotinoids and fipronil and its metabolites in environmental water from coastal bay using disk-based solid-phase extraction and high-performance liquid chromatography–tandem mass spectrometry. Chemosphere 2019, 234, 224–231. [Google Scholar] [PubMed]
- Mahai, G.; Wan, Y.; Xia, W.; Yang, S.; He, Z.; Xu, S. Neonicotinoid insecticides in surface water from the central Yangtze River, China. Chemosphere 2019, 229, 452–460. [Google Scholar]
- Anderson, T.A.; Salice, C.J.; Erickson, R.A.; Mcmurry, S.T.; Cox, S.B.; Smith, L.M. Effects of landuse and precipitation on pesticides and water quality in playa lakes of the southern high plains. Chemosphere 2013, 92, 84–90. [Google Scholar]
- Ruus, A.; Allan, I.J.; Oxnevad, S.; Schaanning, M.T.; Borga, K.; Bakke, T.; Naes, K. In vivo bioaccumulation of contaminants from historically polluted sediments—Relation to bioavailability estimates. Sci. Total Environ. 2013, 442, 336–343. [Google Scholar] [PubMed] [Green Version]
- Donley, N. The USA lags behind other agricultural nations in banning harmful pesticides. Environ. Health 2019, 18, 1–12. [Google Scholar]
- Pan International. Consolidated List of Banned Pesticides. Available online: http://pan-international.org/pan-international-consolidated-list-of-banned-pesticides/ (accessed on 10 October 2020).
- Jeschke, P.; Nauen, R. Neonicotinoids-from zero to hero in insecticide chemistry. Pest Manag. Sci. 2008, 64, 1084–1098. [Google Scholar] [PubMed]
- Sgolastra, F.; Medrzycki, P.; Bortolotti, L.; Maini, S.; Porrini, C.; Simon-Delso, N.; Bosch, J. Bees and pesticide regulation: Lessons from the neonicotinoid experience. Biol. Conserv. 2020, 241, 108356. [Google Scholar]
- Auteri, D.; Arena, M.; Barmaz, S.; Ippolito, A.; Linguadoca, A.; Molnar, T.; Sharp, R.; Szentes, C.; Vagenende, B.; Verani, A. Neonicotinoids and bees: The case of the European regulatory risk assessment. Sci. Total Environ. 2017, 579, 966–971. [Google Scholar] [PubMed]
- European Commission. Current Status of the Neonicotinoids in the EU. Available online: https://ec.europa.eu/food/plant/pesticides/approval_active_substances/approval_renewal/neonicotinoids_en (accessed on 21 October 2020).
- Government of Canada. Update on the Neonicotinoid Pesticides. Health Canada’s Pest Management Regulatory Agency. 2020. Available online: https://www.canada.ca/en/health-canada/services/consumer-product-safety/reports-publications/pesticides-pest-management/fact-sheets-other-resources/update-neonicotinoid-pesticides-january-2020.html (accessed on 8 October 2020).
- Naidu, J. Paraquat, Imidacloprid Pesticide To Be Banned From January 2020. FIJI Sun. 11 October 2019. Available online: http://fijisun.com.fj/2019/10/11/paraquat-imidacloprid-pesticide-to-be-banned-from-january-2020/ (accessed on 21 January 2021).
- Sánchez-Bayo, F.; Goka, K.; Hayasaka, D. Contamination of the aquatic environment with neonicotinoids and its implication for ecosystems. Front. Environ. Sci. 2016, 4, 71. [Google Scholar]
- Goulson, D. Review: An overview of the environmental risks posed by neonicotinoid insecticides. J. Appl. Ecol. 2013, 50, 977–987. [Google Scholar]
- Pisa, L.W.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Downs, C.A.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; Mcfield, M.; et al. Effects of neonicotinoids and fipronil on non-target invertebrates. Environ. Sci. Pollut. Res. 2015, 22, 68–102. [Google Scholar]
- Kreutzweiser, D.P.; Good, K.P.; Chartrand, D.T.; Scarr, T.A.; Thompson, D.G. Are leaves that fall from imidacloprid-treated maple trees to control Asian Longhorned beetles toxic to non-target decomposer organisms? J. Environ. Qual. 2008, 37, 639–646. [Google Scholar]
- Bonmatin, J.-M.; Noome, D.A.; Moreno, H.; Mitchell, E.a.D.; Glauser, G.; Soumana, O.S.; Bijleveld Van Lexmond, M.; Sánchez-Bayo, F. A survey and risk assessment of neonicotinoids in water, soil and sediments of Belize. Environ. Pollut. 2019, 249, 949–958. [Google Scholar]
- Van Dijk, T.C.; Van Staalduinen, M.A.; Van Der Sluijs, J.P. Macro-invertebrate decline in surface water polluted with imidacloprid. PLoS ONE 2013, 8, e62374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrissey, C.A.; Mineau, P.; Devries, J.H.; Sanchez-Bayo, F.; Liess, M.; Cavallaro, M.C.; Liber, K. Neonicotinoid contamination of global surface waters and associated risk to aquatic invertebrates: A review. Environ. Int. 2015, 74, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Douglas, M.R.; Rohr, J.R.; Tooker, J.F. Neonicotinoid insecticide travels through a soil food chain, disrupting biological control of non-target pests and decreasing soya bean yield. J. Appl. Ecol. 2015, 52, 250–260. [Google Scholar] [CrossRef]
- Ewere, E.E.; Powell, D.; Rudd, D.; Reichelt-Brushett, A.; Mouatt, P.; Voelcker, N.H.; Benkendorff, K. Uptake, depuration and sublethal effects of the neonicotinoid, imidacloprid, exposure in Sydney rock oysters. Chemosphere 2019, 230, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ewere, E.E.; Reichelt-Brushett, A.; Benkendorff, K. Imidacloprid and formulated product impacts the fatty acids and enzymatic activities in tissues of Sydney rock oysters, Saccostrea glomerata. Mar. Environ. Res. 2019, 151, 104765. [Google Scholar] [CrossRef]
- Stara, A.; Pagano, M.; Capillo, G.; Fabrello, J.; Sandova, M.; Vazzana, I.; Zuskova, E.; Velisek, J.; Matozzo, V.; Faggio, C. Assessing the effects of neonicotinoid insecticide on the bivalve mollusc Mytilus galloprovincialis. Sci. Total Environ. 2020, 700, 134914. [Google Scholar] [CrossRef]
- Hong, Y.; Huang, Y.; Wu, S.; Yang, X.; Dong, Y.; Xu, D.; Huang, Z. Effects of imidacloprid on the oxidative stress, detoxification and gut microbiota of Chinese mitten crab, Eriocheir sinensis. Sci. Total Environ. 2020, 729, 138276. [Google Scholar] [CrossRef]
- Cimino, A.M.; Boyles, A.L.; Thayer, K.A.; Perry, M.J. Effects of neonicotinoid pesticide exposure on human health: A systematic review. Environ. Health Perspect. 2016, 125, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Ewere, E.E.; Reichelt-Brushett, A.; Benkendorff, K. The neonicotinoid insecticide imidacloprid, but not salinity, impacts the immune system of Sydney rock oyster, Saccostrea glomerata. Sci. Total Environ. 2020, 742, 140538. [Google Scholar] [CrossRef]
- FAO. FAO Yearbook: Fishery and Aquaculture Statistics. Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/fishery/statistics/global-aquaculture-production/en (accessed on 20 September 2020).
- Beck, M.W.; Brumbaugh, R.D.; Airoldi, L.; Carranza, A.; Coen, L.D.; Crawford, C.; Defeo, O.; Edgar, G.J.; Hancock, B.; Kay, M.C. Oyster reefs at risk and recommendations for conservation, restoration, and management. Bioscience 2011, 61, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Peng, M.; Oleson, K.L. Beach recreationalists’ willingness to pay and economic implications of coastal water quality problems in Hawaii. Ecol Econom 2017, 136, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Mcleod, I.M.; Zu Ermgassen, P.S.E.; Gillies, C.L.; Hancock, B.; Humphries, A. Can Bivalve Habitat Restoration Improve Degraded Estuaries? In Coasts and Estuaries; Wolanski, E., Day, J.W., Elliott, M., Ramachandran, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 427–442. [Google Scholar]
- Tomizawa, M.; Yamamoto, I. Structure-activity relationships of nicotinoids and imidacloprid analogs. J. Pestic. Sci. 1993, 18, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Jeschke, P.; Nauen, R.; Schindler, M.; Elbert, A. Overview of the status and global strategy for neonicotinoids. J. Agric. Food Chem. 2011, 59, 2897–2908. [Google Scholar] [CrossRef] [PubMed]
- Elbert, A.; Haas, M.; Springer, B.; Thielert, W.; Nauen, R. Mini-review Applied aspects of neonicotinoid uses in crop protection. Pest Manag. Sci. 2008, 64, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.A.; Chang, C.-H.; Tao, L.; Chen, M. Distributions of neonicotinoid insecticides in the Commonwealth of Massachusetts: A temporal and spatial variation analysis for pollen and honey samples. Environ. Chem. 2015, 13, 4–11. [Google Scholar] [CrossRef]
- Fauser, A.; Sandrock, C.; Neumann, P.; Sadd, B.M. Neonicotinoids override a parasite exposure impact on hibernation success of a key bumblebee pollinator. Ecol. Entomol. 2017, 42, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Phugare, S.S.; Kalyani, D.C.; Gaikwad, Y.B.; Jadhav, J.P. Microbial degradation of imidacloprid and toxicological analysis of its biodegradation metabolites in silkworm (Bombyx mori). Chem. Eng. J. 2013, 230, 27–35. [Google Scholar] [CrossRef]
- Chiovarou, E.D.; Siewicki, T.C. Comparison of storm intensity and application timing on modeled transport and fate of six contaminants. Sci. Total Environ. 2008, 389, 87–100. [Google Scholar] [CrossRef]
- Leblanc, H.M.K.; Culp, J.M.; Baird, D.J.; Alexander, A.C.; Cessna, A.J. Single versus combined lethal effects of three agricultural insecticides on larvae of the freshwater insect Chironomus dilutus. Arch. Environ. Contam. Toxicol. 2012, 63, 378–390. [Google Scholar] [CrossRef]
- Moertl, M.; Vehovszky, A.; Klatyik, S.; Takacs, E.; Gyori, J.; Szekacs, A. Neonicotinoids: Spreading, Translocation and Aquatic Toxicity. Int. J. Environ. Res. Public Health 2020, 17, 2006. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, K.J.A.; Santana, M.B.; Do Nascimento, J.L.; Picanco-Diniz, D.L.; Maues, L.A.; Santos, S.N.; Ferreira, V.M.; Alfonso, M.; Duran, R.; Faro, L.R. Behavioral and biochemical effects of neonicotinoid thiamethoxam on the cholinergic system in rats. Ecotoxicol. Environ. Saf. 2010, 73, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Tomizawa, M.; Casida, J.E. Neonicotinoid insecticide toxicology: Mechanisms of selective action. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 247–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon-Delso, N.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Chagnon, M.; Downs, C.; Furlan, L.; Gibbons, D.W.; Giorio, C.; Girolami, V.; et al. Systemic insecticides (neonicotinoids and fipronil): Trends, uses, mode of action and metabolites. Environ. Sci. Pollut. Res. 2015, 22, 5–34. [Google Scholar] [CrossRef] [PubMed]
- Thany, S.H. Neonicotinoid Insecticides Historical Evolution and Resistance Mechanisms. In Insect Nicotinic Acetylcholine Receptors; Thany, S.H., Ed.; Springer: Cham, Switzerland, 2010; pp. 75–83. [Google Scholar]
- Pagano, M.; Stara, A.; Aliko, V.; Faggio, C. Impact of neonicotinoids to aquatic invertebrates—in vitro studies on Mytilus galloprovincialis: A review. J. Mar. Sci. Eng. 2020, 8, 801. [Google Scholar] [CrossRef]
- Cavallaro, M.C.; Morrissey, C.A.; Headley, J.V.; Peru, K.M.; Liber, K. Comparative chronic toxicity of imidacloprid, clothianidin, and thiamethoxam to Chironomus dilutus and estimation of toxic equivalency factors. Environ. Toxicol. Chem. 2017, 36, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Tomizawa, M.; Maltby, D.; Talley, T.T.; Durkin, K.A.; Medzihradszky, K.F.; Burlingame, A.L.; Taylor, P.; Casida, J.E. Atypical nicotinic agonist bound conformations conferring subtype selectivity. Proc. Natl. Acad. Sci. USA 2008, 105, 1728–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United States Environmental Protection Agency. Schedule for Review of Neonicotinoid Pesticides. Available online: https://www.epa.gov/pollinator-protection/schedule-review-neonicotinoid-pesticides (accessed on 2 September 2020).
- Anderson, J.C.; Dubetz, C.; Palace, V.P. Neonicotinoids in the Canadian aquatic environment: A literature review on current use products with a focus on fate, exposure, and biological effects. Sci. Total Environ. 2015, 505, 409–422. [Google Scholar] [CrossRef]
- Bonmatin, J.M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.a.D.; et al. Environmental fate and exposure; neonicotinoids and fipronil. Environ. Sci. Pollut. Res. 2015, 22, 35–67. [Google Scholar] [CrossRef]
- Wood, T.J.; Goulson, D. The environmental risks of neonicotinoid pesticides: A review of the evidence post 2013. Environ. Sci. Pollut. Res. 2017, 24, 17285–17325. [Google Scholar] [CrossRef]
- Main, A.R.; Headley, J.V.; Peru, K.M.; Michel, N.L.; Cessna, A.J. Widespread Use and Frequent Detection of Neonicotinoid Insecticides in Wetlands of Canada’s Prairie Pothole Region. PLoS ONE 2014, 9, e92821. [Google Scholar] [CrossRef]
- Kennedy, J.M.; Talbert, R.E. Comparative persistence of dinitroaniline type herbicides on the soil surface. Weed Sci. 1977, 25, 373–381. [Google Scholar] [CrossRef]
- El-Hamady, S.E.; Kubiak, R.; Derbalah, A.S. Fate of imidacloprid in soil and plant after application to cotton seeds. Chemosphere 2008, 71, 2173–2179. [Google Scholar] [CrossRef] [PubMed]
- Thuyet, D.Q.; Watanabe, H.; Yamazaki, K.; Takagi, K. Photodegradation of imidacloprid and fipronil in rice–paddy water. Bull. Environ. Contam. Toxicol. 2011, 86, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Segawa, R.; Walters, J. Preliminary Monitoring Results of Imidacloprid and Cyfluthrin Applications for Glassy-Winged Sharpshooter Control in Residential Areas of Santa Clara County; California Department of Pesticide Regulation: Sacramento, CA, USA, 2002.
- Segawa, R.; Walters, J.; Fan, S. Preliminary Monitoring Results of Imidacloprid and Cyfluthrin Applications for Glassy-Winged Sharpshooter Control in a Residential Area of Solano County; California Department of Pesticide Regulation: Sacramento, CA, USA, 2004.
- Segawa, R.; Walters, J.; Fan, S.; Matsumoto, S.; Casjens, H. Preliminary Monitoring Results of Imidacloprid and Cyfluthrin Applications for Glassy-Winged Sharpshooter Control in Commercial Areas of Imperial County; California Department of Pesticide Regulation: Sacramento, CA, USA, 2002.
- Walters, J.; Casjens, H.; Goh, K.S. Preliminary Monitoring Results of Imidacloprid and Cyfluthrin Applications for Glassy-Winged Sharpshooter Control in Commercial Areas of Butte County; Regulation, D.O.P., Ed.; California Department of Pesticide Regulation: Sacramento, CA, USA, 2001; pp. 1–5.
- Girolami, V.; Marzaro, M.; Vivan, L.; Mazzon, L.; Greatti, M.; Giorio, C.; Marton, D.; Tapparo, A. Fatal powdering of bees in flight with particulates of neonicotinoids seed coating and humidity implication. J. Appl Entomol 2012, 136, 17–26. [Google Scholar] [CrossRef]
- Pochi, D.; Biocca, M.; Fanigliulo, R.; Pulcini, P.; Conte, E. Potential exposure of bees, Apis mellifera L., to particulate matter and pesticides derived from seed dressing during maize sowing. Bull. Environ. Contam. Toxicol. 2012, 89, 354–361. [Google Scholar] [CrossRef]
- Greatti, M.; Sabatini, G.A.; Barbattini, R.; Rossi, S.; Stravisi, A. Risk of environmental contamination by the active ingredient imidacloprid used for corn seed dressing. Preliminary results. Bull. Insectol. 2003, 56, 69–72. [Google Scholar]
- Krupke, C.H.; Hunt, G.J.; Eitzer, B.D.; Andino, G.; Given, K. Multiple Routes of Pesticide Exposure for Honey Bees Living Near Agricultural Fields. PLoS ONE 2012, 7, e29268. [Google Scholar] [CrossRef]
- Biocca, M.; Fanigliulo, R.; Gallo, P.; Pulcini, P.; Perrino, C.; Pochi, D. Assessing dust drift from dressed seeds by air sampling. Asp. Appl. Biol. 2014, 122, 103–111. [Google Scholar]
- Forero, L.G.; Limay-Rios, V.; Xue, Y.; Schaafsma, A. Concentration and movement of neonicotinoids as particulate matter downwind during agricultural practices using air samplers in southwestern Ontario, Canada. Chemosphere 2017, 188, 130–138. [Google Scholar] [CrossRef]
- Selim, H.M.; Jeong, C.Y.; Elbana, T.A. Transport of imidacloprid in soils: Miscible displacement experiments. Soil Sci. 2010, 175, 375–381. [Google Scholar] [CrossRef]
- Horwood, M.A. Rapid degradation of termiticides under field conditions. Aust. J. Entomol. 2007, 46, 75–78. [Google Scholar] [CrossRef]
- Fratini, S.; Vigiani, V.; Vannini, M.; Cannicci, S. Terebralia palustris (Gastropoda; Potamididae) in a Kenyan mangal: Size structure, distribution and impact on the consumption of leaf litter. Mar. Biol. 2004, 144, 1173–1182. [Google Scholar] [CrossRef]
- Pang, S.; Lin, Z.; Zhang, Y.; Zhang, W.; Alansary, N.; Mishra, S.; Bhatt, P.; Chen, S. Insights into the Toxicity and Degradation Mechanisms of Imidacloprid Via Physicochemical and Microbial Approaches. Toxics 2020, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Anhalt, J.C.; Moorman, T.B.; Koskinen, W.C. Biodegradation of imidacloprid by an isolated soil microorganism. J. Environ. Sci. Health B 2007, 42, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.; Dorrian, S.J.; Russell, R.J.; Oakeshott, J.G. Biotransformation of the neonicotinoid insecticides imidacloprid and thiamethoxam by Pseudomonas sp. 1G. Biochem. Biophys. Res. Commun. 2009, 380, 710–714. [Google Scholar] [CrossRef]
- Guo, L.; Dai, Z.; Guo, J.; Yang, W.; Ge, F.; Dai, Y. Oligotrophic bacterium Hymenobacter latericoloratus CGMCC 16346 degrades the neonicotinoid imidacloprid in surface water. AMB Express 2020, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zhan, H.; Wang, H.; Liao, L.; Feng, Y.; Fan, X.; Zhang, L.; Chen, S. Kinetics and novel degradation pathway of permethrin in Acinetobacter baumannii ZH-14. Front. Microbiol. 2018, 9, 98. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Singh, B.; Gupta, V. Assessment of imidacloprid degradation by soil-isolated Bacillus alkalinitrilicus. Environ. Monit. Assess. 2014, 186, 7183–7193. [Google Scholar] [CrossRef]
- Júnior, R.P.S.; Smelt, J.H.; Boesten, J.J.; Hendriks, R.F.A.; van der Zee, S.E. Preferential flow of bromide, bentazon, and imidacloprid in a Dutch clay soil. J. Environ. Qual. 2004, 33, 1473–1486. [Google Scholar]
- Tisler, T.; Jemec, A.; Mozetic, B.; Trebse, P. Hazard identification of imidacloprid to aquatic environment. Chemosphere 2009, 76, 907–914. [Google Scholar] [CrossRef]
- Lu, Z.; Challis, J.K.; Wong, C.S. Quantum yields for direct photolysis of neonicotinoid insecticides in water: Implications for exposure to nontarget aquatic organisms. Environ. Sci. Technol. Lett. 2015, 2, 188–192. [Google Scholar] [CrossRef]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pestic Biochem Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, P.K.S. Use of biomarkers in environmental monitoring. Ocean Coast. Manag. 2009, 52, 348–354. [Google Scholar] [CrossRef]
- Lam, P.K.; Gray, J.S. The use of biomarkers in environmental monitoring programmes. Mar. Pollut. Bull. 2003, 46, 182–186. [Google Scholar] [CrossRef]
- Shan, Y.; Yan, S.; Hong, X.; Zha, J.; Qin, J. Effect of imidacloprid on the behavior, antioxidant system, multixenobiotic resistance, and histopathology of Asian freshwater clams (Corbicula fluminea). Aquat. Toxicol. 2020, 218, 105333. [Google Scholar] [CrossRef]
- Smina, A.H.; Samira, B.; Mohamed, D.; Houria, B. Evaluation of acetylcholinesterase, glutathione S-transferase and catalase activities in the land snail Helix aspersa exposed to thiamethoxam. J. Ent. Zool. 2016, 4, 369–374. [Google Scholar]
- Hamlet, S.A.; Bensoltane, S.; Djekoun, M.; Yassi, F.; Berrebbah, H. Histological changes and biochemical parameters in the hepatopancreas of terrestrial gastropod Helix aspersa as biomarkers of neonicotinoid insecticide exposure. Afr. J. Biotechnol. 2012, 11, 16277–16283. [Google Scholar]
- Kuchovská, E.; Morin, B.; López-Cabeza, R.; Barré, M.; Gouffier, C.; Bláhová, L.; Cachot, J.; Bláha, L.; Gonzalez, P. Comparison of imidacloprid, propiconazole, and nanopropiconazole effects on the development, behavior, and gene expression biomarkers of the Pacific oyster (Magallana gigas). Sci. Total Environ. 2020, 142921. [Google Scholar] [CrossRef]
- Simms, L.C.; Ester, A.; Wilson, M.J. Control of slug damage to oilseed rape and wheat with imidacloprid seed dressings in laboratory and field experiments. Crop Prot. 2006, 25, 549–555. [Google Scholar] [CrossRef]
- Chmist, J.; Szoszkiewicz, K.; Drożdżyński, D. Behavioural Responses of Unio tumidus Freshwater Mussels to Pesticide Contamination. Arch. Environ. Contam. Toxicol. 2019, 77, 432–442. [Google Scholar] [CrossRef] [Green Version]
- Oliver, R.; Fuhrmann, M.; Hick, P. Effect of air exposure, handling stress and imidacloprid on the susceptibility of Crassostrea gigas to Ostreid herpesvirus 1 (OsHV-1). Aquac. Environ. Interact. 2019, 11, 685–699. [Google Scholar] [CrossRef]
- Sawasdee, B.; Köhler, H.-R. Embryo toxicity of pesticides and heavy metals to the ramshorn snail, Marisa cornuarietis (Prosobranchia). Chemosphere 2009, 75, 1539–1547. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, M.C.; Baxter, L.R.; Maul, J.D.; Hanson, M.L.; Hoekstra, P.F. Comprehensive characterization of the acute and chronic toxicity of the neonicotinoid insecticide thiamethoxam to a suite of aquatic primary producers, invertebrates, and fish. Environ. Toxicol. Chem. 2017, 36, 2838–2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prosser, R.S.; De Solla, S.R.; Holman, E.A.; Osborne, R.; Robinson, S.A.; Bartlett, A.J.; Maisonneuve, F.J.; Gillis, P.L. Sensitivity of the early-life stages of freshwater mollusks to neonicotinoid and butenolide insecticides. Environ. Pollut. 2016, 218, 428–435. [Google Scholar] [CrossRef]
- Hamlet, S.A.; Djekoun, M.; Smati, M.; Semassel, A.; Bensoltane, S.D.; Berrebbah, H. Histopathological effects of neonicotinoid insecticide in the hepatopancreas of terrestrial gastropod Helix aspersa. Fresenius Environ. Bull. 2014, 23, 3041–3047. [Google Scholar]
- Patil, A. Thiamethoxam induced histopathological alterations in hepatopancreas of freshwater bivalve Parreysia cylindrica. J. Entomol. Zool. Stud. 2019, 7, 1236–1240. [Google Scholar]
- Tufi, S.; Stel, J.M.; De Boer, J.; Lamoree, M.H.; Leonards, P.E.G. Metabolomics to Explore Imidacloprid-Induced Toxicity in the Central Nervous System of the Freshwater Snail Lymnaea stagnalis. Environ. Sci. Technol. 2015, 49, 14529–14536. [Google Scholar] [CrossRef]
- Tomizawa, M.; Casida, J.E. Unique Neonicotinoid Binding Conformations Conferring Selective Receptor Interactions. J. Agric. Food Chem. 2011, 59, 2825–2828. [Google Scholar] [CrossRef]
- Vehovszky, Á.; Farkas, A.; Ács, A.; Stoliar, O.; Székács, A.; Mörtl, M.; Győri, J. Neonicotinoid insecticides inhibit cholinergic neurotransmission in a molluscan (Lymnaea stagnalis) nervous system. Aquat. Toxicol. 2015, 167, 172–179. [Google Scholar] [CrossRef] [Green Version]
- Dondero, F.; Negri, A.; Boatti, L.; Marsano, F.; Mignone, F.; Viarengo, A. Transcriptomic and proteomic effects of a neonicotinoid insecticide mixture in the marine mussel (Mytilus galloprovincialis, Lam.). Sci. Total Environ. 2010, 408, 3775–3786. [Google Scholar] [CrossRef]
- Cossi, P.F.; Herbert, L.T.; Yusseppone, M.S.; Pérez, A.F.; Kristoff, G. Toxicity evaluation of the active ingredient acetamiprid and a commercial formulation (Assail® 70) on the non-target gastropod Biomphalaria straminea (Mollusca: Planorbidae). Ecotoxicol. Environ. Saf. 2020, 192, 110248. [Google Scholar] [CrossRef] [PubMed]
- Moncaleano-Niño, A.M.; Luna-Acosta, A.; Gómez-Cubillos, M.C.; Villamil, L.; Ahrens, M.J. Cholinesterase activity in the cup oyster Saccostrea sp. exposed to chlorpyrifos, imidacloprid, cadmium and copper. Ecotoxicol. Environ. Saf. 2018, 151, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Sumon, K.A.; Ritika, A.K.; Peeters, E.T.H.M.; Rashid, H.; Bosma, R.H.; Rahman, M.S.; Fatema, M.K.; Van Den Brink, P.J. Effects of imidacloprid on the ecology of sub-tropical freshwater microcosms. Environ. Pollut. 2018, 236, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Rico, A.; Arenas-Sánchez, A.; Pasqualini, J.; García-Astillero, A.; Cherta, L.; Nozal, L.; Vighi, M. Effects of imidacloprid and a neonicotinoid mixture on aquatic invertebrate communities under Mediterranean conditions. Aquat. Toxicol. 2018, 204, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, M.C.; Emburey, S.; Hommen, U.; Baxter, L.R.; Hoekstra, P.F.; Hanson, M.L.; Thompson, H.; Hamer, M. A freshwater mesocosm study into the effects of the neonicotinoid insecticide thiamethoxam at multiple trophic levels. Environ. Pollut. 2018, 242, 1444–1457. [Google Scholar] [CrossRef] [PubMed]
- Salerno, J.; Bennett, C.J.; Holman, E.; Gillis, P.L.; Sibley, P.K.; Prosser, R.S. Sensitivity of multiple life stages of 2 freshwater mussel species (Unionidae) to various pesticides detected in Ontario (Canada) surface waters. Environ. Toxicol. Chem. 2018, 37, 2871–2880. [Google Scholar] [CrossRef] [PubMed]
- Miles, J.C.; Hua, J.; Sepulveda, M.S.; Krupke, C.H.; Hoverman, J.T. Effects of clothianidin on aquatic communities: Evaluating the impacts of lethal and sublethal exposure to neonicotinoids. PLoS ONE 2017, 12, e0174171. [Google Scholar] [CrossRef]
- Colombo, V.; Mohr, S.; Berghahn, R.; Pettigrove, V.J. Structural Changes in a Macrozoobenthos Assemblage After Imidacloprid Pulses in Aquatic Field-Based Microcosms. Arch. Environ. Contam. Toxicol. 2013, 65, 683–692. [Google Scholar] [CrossRef]
- Gupta, S.; Gajbhiye, V.; Gupta, R. Effect of light on the degradation of two neonicotinoids viz acetamiprid and thiacloprid in soil. Bull. Environ. Contam. Toxicol. 2008, 81, 185–189. [Google Scholar] [CrossRef]
- Gupta, S.K.; Singh, J. Evaluation of mollusc as sensitive indicator of heavy metal pollution in aquatic system: A review. IIOAB J. 2011, 2, 49–57. [Google Scholar]
- Marigómez, I.; Soto, M.; Cajaraville, M.P.; Angulo, E.; Giamberini, L. Cellular and subcellular distribution of metals in molluscs. Microsc. Res. Tech. 2002, 56, 358–392. [Google Scholar] [CrossRef] [PubMed]
- Zorita, I.; Larreta, J.; Montero, N.; Rodríguez, J.G.; Franco, J.; Borja, Á. Evaluation of the use of bioaccumulation and biological effects tools in caged mussels, within the European water framework directive. Chem. Ecol. 2015, 31, 432–445. [Google Scholar] [CrossRef]
- Jacomini, A.E.; Avelar, W.E.P.; Martinêz, A.S.; Bonato, P.S. Bioaccumulation of atrazine in freshwater bivalves Anodontites trapesialis (Lamarck, 1819) and Corbicula fluminea (Müller, 1774). Arch. Environ. Contam. Toxicol. 2006, 51, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Beltran, K.S.; Pocsidio, G.N. Acetylcholinesterase activity in Corbicula fluminea Mull., as a biomarker of organophosphate pesticide pollution in Pinacanauan River, Philippines. Environ. Monit. Assess. 2010, 165, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Beyer, J.; Green, N.W.; Brooks, S.; Allan, I.J.; Ruus, A.; Gomes, T.; Bråte, I.L.N.; Schøyen, M. Blue mussels (Mytilus edulis spp.) as sentinel organisms in coastal pollution monitoring: A review. Mar. Environ. Res. 2017, 130, 338–365. [Google Scholar] [CrossRef]
- Dawson, A.; Huston, W.; Kawaguchi, S.; King, C.; Cropp, R.; Wild, S.; Eisenmann, P.; Townsend, K.; Bengtson Nash, S. Uptake and Depuration Kinetics Influence Microplastic Bioaccumulation and Toxicity in Antarctic Krill (Euphausia superba). Environ. Sci. Technol. 2018, 52, 3195–3201. [Google Scholar] [CrossRef] [PubMed]
- Berlioz-Barbier, A.; Buleté, A.; Faburé, J.; Garric, J.; Cren-Olivé, C.; Vulliet, E. Multi-residue analysis of emerging pollutants in benthic invertebrates by modified micro-quick-easy-cheap-efficient-rugged-safe extraction and nanoliquid chromatography–nanospray–tandem mass spectrometry analysis. J. Chromatogr. A 2014, 1367, 16–32. [Google Scholar] [CrossRef]
- Mineau, P.; Palmer, C. The impact of the nation’s most widely used insecticides on birds: Neonicotinoid insecticides and birds. American Bird Conservancy, Washington, DC. Available online: https://extension.entm.purdue.edu/neonicotinoids/PDF/TheImpactoftheNationsMostWidelyUsedInsecticidesonBirds.pdf (accessed on 20 January 2020).
- Ford, K.A.; Casida, J.E. Chloropyridinyl neonicotinoid insecticides: Diverse molecular substituents contribute to facile metabolism in mice. Chem. Res. Toxicol. 2006, 19, 944–951. [Google Scholar] [CrossRef]
- Kimura-Kuroda, J.; Komuta, Y.; Kuroda, Y.; Hayashi, M.; Kawano, H. Nicotine-like effects of the neonicotinoid insecticides acetamiprid and imidacloprid on cerebellar neurons from neonatal rats. PLoS ONE 2012, 7, e32432. [Google Scholar] [CrossRef]
- Stara, A.; Bellinvia, R.; Velisek, J.; Strouhova, A.; Kouba, A.; Faggio, C. Acute exposure of common yabby (Cherax destructor) to the neonicotinoid pesticide. Sci. Total Environ. 2019, 665, 718–723. [Google Scholar] [CrossRef]
- El-Gendy, K.S.; Radwan, M.A.; Gad, A.F.; Khamis, A.E.; Eshra, E.H. Use of multiple endpoints to investigate the ecotoxicological effects of abamectin and thiamethoxam on Theba pisana snails. Ecotoxicol. Environ. Saf. 2019, 167, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Archambault, J.M.; Bergeron, C.M.; Cope, W.G.; Richardson, R.J.; Heilman, M.A.; Corey Iii, J.E.; Netherland, M.D.; Heise, R.J. Sensitivity of freshwater molluscs to hydrilla-targeting herbicides: Providing context for invasive aquatic weed control in diverse ecosystems. J. Freshw. Ecol. 2015, 30, 335–348. [Google Scholar] [CrossRef]
- Zhang, H.; Hong, X.; Yan, S.; Zha, J.; Qin, J. Environmentally relevant concentrations of bifenthrin induce changes in behaviour, biomarkers, histological characteristics, and the transcriptome in Corbicula fluminea. Sci. Total Environ. 2020, 728, 138821. [Google Scholar] [CrossRef] [PubMed]
- Hazelton, P.D.; Du, B.; Haddad, S.P.; Fritts, A.K.; Chambliss, C.K.; Brooks, B.W.; Bringolf, R.B. Chronic fluoxetine exposure alters movement and burrowing in adult freshwater mussels. Aquat. Toxicol. 2014, 151, 27–35. [Google Scholar] [CrossRef]
- Cope, W.G.; Bringolf, R.B.; Buchwalter, D.B.; Newton, T.J.; Ingersoll, C.G.; Wang, N.; Augspurger, T.; Dwyer, F.J.; Barnhart, M.C.; Neves, R.J. Differential exposure, duration, and sensitivity of unionoidean bivalve life stages to environmental contaminants. J. N. Am. Benthol. Soc. 2008, 27, 451–462. [Google Scholar] [CrossRef]
- Graney, R.L.; Giesy, J.P., Jr. Alterations in the oxygen consumption, condition index and concentration of free amino acids in Corbicula fluminea,(mollusca: Pelecypoda) exposed to sodium dodecyl sulfate. Environ. Toxicol. Chem. 1988, 7, 301–315. [Google Scholar] [CrossRef]
- Singh, A.; Leppanen, C. Known Target and Nontarget Effects of the Novel Neonicotinoid Cycloxaprid to Arthropods: A Systematic Review. Integr. Environ. Assess. Manag. 2020, 16, 831–840. [Google Scholar] [CrossRef]
- Matozzo, V.; Chinellato, A.; Munari, M.; Finos, L.; Bressan, M.; Marin, M.G. First evidence of immunomodulation in bivalves under seawater acidification and increased temperature. PLoS ONE 2012, 7, e33820. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wang, H.; Chen, H.; Wang, M.; Zhou, Z.; Qiu, L.; Wang, L.; Song, L. The transcriptional response of the Pacific oyster Crassostrea gigas under simultaneous bacterial and heat stresses. Dev. Comp. Immunol. 2019, 94, 1–10. [Google Scholar] [CrossRef]
- Song, L.; Wang, L.; Qiu, L.; Zhang, H. Bivalve Immunity. In Invertebrate Immunity; Söderhäll, K., Ed.; Springer: Boston, MA, USA, 2010; pp. 44–65. [Google Scholar]
- Moreau, P.; Burgeot, T.; Renault, T. Pacific oyster (Crassostrea gigas) hemocyte are not affected by a mixture of pesticides in short-term in vitro assays. Environ. Sci. Pollut. Res. 2014, 21, 4940–4949. [Google Scholar] [CrossRef]
- Altinok, I.; Capkin, E. Histopathology of rainbow trout exposed to sublethal concentrations of methiocarb or endosulfan. Toxicol. Pathol. 2007, 35, 405–410. [Google Scholar] [CrossRef]
- Thompson, D.A.; Lehmler, H.-J.; Kolpin, D.W.; Hladik, M.L.; Vargo, J.D.; Schilling, K.E.; Lefevre, G.H.; Peeples, T.L.; Poch, M.C.; Laduca, L.E. A critical review on the potential impacts of neonicotinoid insecticide use: Current knowledge of environmental fate, toxicity, and implications for human health. Environ. Sci. Processes Impacts 2020, 22, 1315–1346. [Google Scholar] [CrossRef] [PubMed]
- Ensley, S.M. Chapter 48—Neonicotinoids. In Veterinary Toxicology, 2nd ed.; Gupta, R.C., Ed.; Academic Press: Boston, MA, USA, 2012; pp. 596–598. [Google Scholar]
- Silva, C.O.; Novais, S.C.; Alves, L.M.F.; Soares, A.M.V.M.; Barata, C.; Lemos, M.F.L. Linking cholinesterase inhibition with behavioural changes in the sea snail Gibbula umbilicalis: Effects of the organophosphate pesticide chlorpyrifos. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 225, 108570. [Google Scholar] [CrossRef] [PubMed]
- Manduzio, H.; Rocher, B.; Durand, F.; Galap, C.; Leboulenger, F. The point about oxidative stress in molluscs. Invertebr. Surviv. J. 2005, 2, 91–104. [Google Scholar]
- Richardson, B.J.; Mak, E.; De Luca-Abbott, S.B.; Martin, M.; Mcclellan, K.; Lam, P.K. Antioxidant responses to polycyclic aromatic hydrocarbons and organochlorine pesticides in green-lipped mussels (Perna viridis): Do mussels “integrate” biomarker responses? Mar. Pollut. Bull. 2008, 57, 503–514. [Google Scholar] [CrossRef]
- Solé, M. Assessment of the results of chemical analyses combined with the biological effects of organic pollution on mussels. TrAC 2000, 19, 1–9. [Google Scholar] [CrossRef]
- Topal, A.; Alak, G.; Ozkaraca, M.; Yeltekin, A.C.; Comaklı, S.; Acıl, G.; Kokturk, M.; Atamanalp, M. Neurotoxic responses in brain tissues of rainbow trout exposed to imidacloprid pesticide: Assessment of 8-hydroxy-2-deoxyguanosine activity, oxidative stress and acetylcholinesterase activity. Chemosphere 2017, 175, 186–191. [Google Scholar] [CrossRef]
- Shila, S.; Kokilavani, V.; Subathra, M.; Panneerselvam, C. Brain regional responses in antioxidant system to α-lipoic acid in arsenic intoxicated rat. Toxicology 2005, 210, 25–36. [Google Scholar] [CrossRef]
- Desai, S.N.; Farris, F.F.; Ray, S.D. Lipid Peroxidation. In Encyclopedia of Toxicology, 3rd ed.; Wexler, P., Ed.; Academic Press: Oxford, UK, 2014; pp. 89–93. [Google Scholar]
- Barber, B.J.; Blake, N.J. Energy storage and utilization in relation to gametogenesis in Argopecten irradians concentricus (Say). J. Exp. Mar. Biol. Ecol. 1981, 52, 121–134. [Google Scholar] [CrossRef]
- Dondero, F.; Banni, M.; Negri, A.; Boatti, L.; Dagnino, A.; Viarengo, A. Interactions of a pesticide/heavy metal mixture in marine bivalves: A transcriptomic assessment. BMC Genom. 2011, 12, 195. [Google Scholar] [CrossRef] [Green Version]
- Przeslawski, R. A review of the effects of environmental stress on embryonic development within intertidal gastropod egg masses. Molluscan Res. 2004, 24, 43–63. [Google Scholar] [CrossRef]
- Behrens, D.; Rouxel, J.; Burgeot, T.; Akcha, F. Comparative embryotoxicity and genotoxicity of the herbicide diuron and its metabolites in early life stages of Crassostrea gigas: Implication of reactive oxygen species production. Aquat. Toxicol. 2016, 175, 249–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewere, E.E. Impacts of the Neonicotinoid Insecticide, Imidacloprid, on Sydney rock Oyster. Ph.D. Thesis, Southern Cross University, Lismore, NSW, Australia, 2020. [Google Scholar] [CrossRef]




| Knowledge Gap | Further Research |
|---|---|
| No data on the impact of neonicotinoids on gametes or fertilisation success of molluscs. | Acute studies to identify the impacts of neonicotinoids to gametes and fertilisation success of molluscs, as well as acute and chronic trials on the development processes and metamorphosis of early life stages. For species that undergo external fertilisation (e.g., bivalves), there is also the need for studies determining the toxicity of neonicotinoids to sperms under natural environmental conditions. |
| Minimal data on the impacts of neonicotinoids on the safety and nutritional quality of edible molluscs. | Studies determining the acute and chronic effects of neonicotinoids on the safety and nutritional quality of edible species of molluscs, especially commercial species of gastropods and bivalves. |
| No data on the impacts of neonicotinoids on some ecologically important classes of molluscs. | Studies to determine the lethal and sublethal impacts of neonicotinoids to all the classes of molluscs, including classes that are not economically important. |
| Limited data on the impact of neonicotinoids mixtures on molluscs | Studies to identify the possible synergistic or antagonistic impacts of neonicotinoid mixtures as well as in combination with other chemicals and/or other environmental stressors on molluscs. |
| Limited data on the accumulation and elimination potential of neonicotinoids in molluscs. | More exposure experiments to determine the rate of accumulation, metabolisms and depuration of neonicotinoids in molluscs, to understand the risk of the possible exposure of other organisms higher along the food chain. |
| Very limited data exist on the impacts of neonicotinoids under stressful environmental regimes in molluscs. | Studies determining the effects of neonicotinoids on molluscs under various conditions, including salinity, temperature and pH. |
| Very limited data on the impacts of acute and chronic neonicotinoids exposure in molluscs under natural conditions | Mesocosms and field experiments to determine the impacts of neonicotinoids exposure to molluscs under natural conditions. |
| Very limited data exists on the genetic changes and regulatory mechanisms underlying molluscs response to neonicotinoids | Transcriptomics, DNA methylation and targeted gene expression studies to assess the physiological response of molluscs to neonicotinoids. |
| Limited data exist on the impacts of neonicotinoids on the physiology and immune system of molluscs. | Controlled manipulative studies to establish the causal effects on physiological and immunological responses of molluscs to neonicotinoids and any consequent tertiary effects on disease resistance, growth and mortality. |
| No data on the possible carry-over effects to the offspring due to adult exposure to neonicotinoids | Manipulative experiments to determine transgenerational impacts or resistance in offspring of molluscs that have been exposed to neonicotinoids. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ewere, E.E.; Reichelt-Brushett, A.; Benkendorff, K. Impacts of Neonicotinoids on Molluscs: What We Know and What We Need to Know. Toxics 2021, 9, 21. https://doi.org/10.3390/toxics9020021
Ewere EE, Reichelt-Brushett A, Benkendorff K. Impacts of Neonicotinoids on Molluscs: What We Know and What We Need to Know. Toxics. 2021; 9(2):21. https://doi.org/10.3390/toxics9020021
Chicago/Turabian StyleEwere, Endurance E, Amanda Reichelt-Brushett, and Kirsten Benkendorff. 2021. "Impacts of Neonicotinoids on Molluscs: What We Know and What We Need to Know" Toxics 9, no. 2: 21. https://doi.org/10.3390/toxics9020021
APA StyleEwere, E. E., Reichelt-Brushett, A., & Benkendorff, K. (2021). Impacts of Neonicotinoids on Molluscs: What We Know and What We Need to Know. Toxics, 9(2), 21. https://doi.org/10.3390/toxics9020021