
Biomanufacturing Biotinylated Magnetic Nanomaterial via Construction and Fermentation of Genetically Engineered Magnetotactic Bacteria

 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
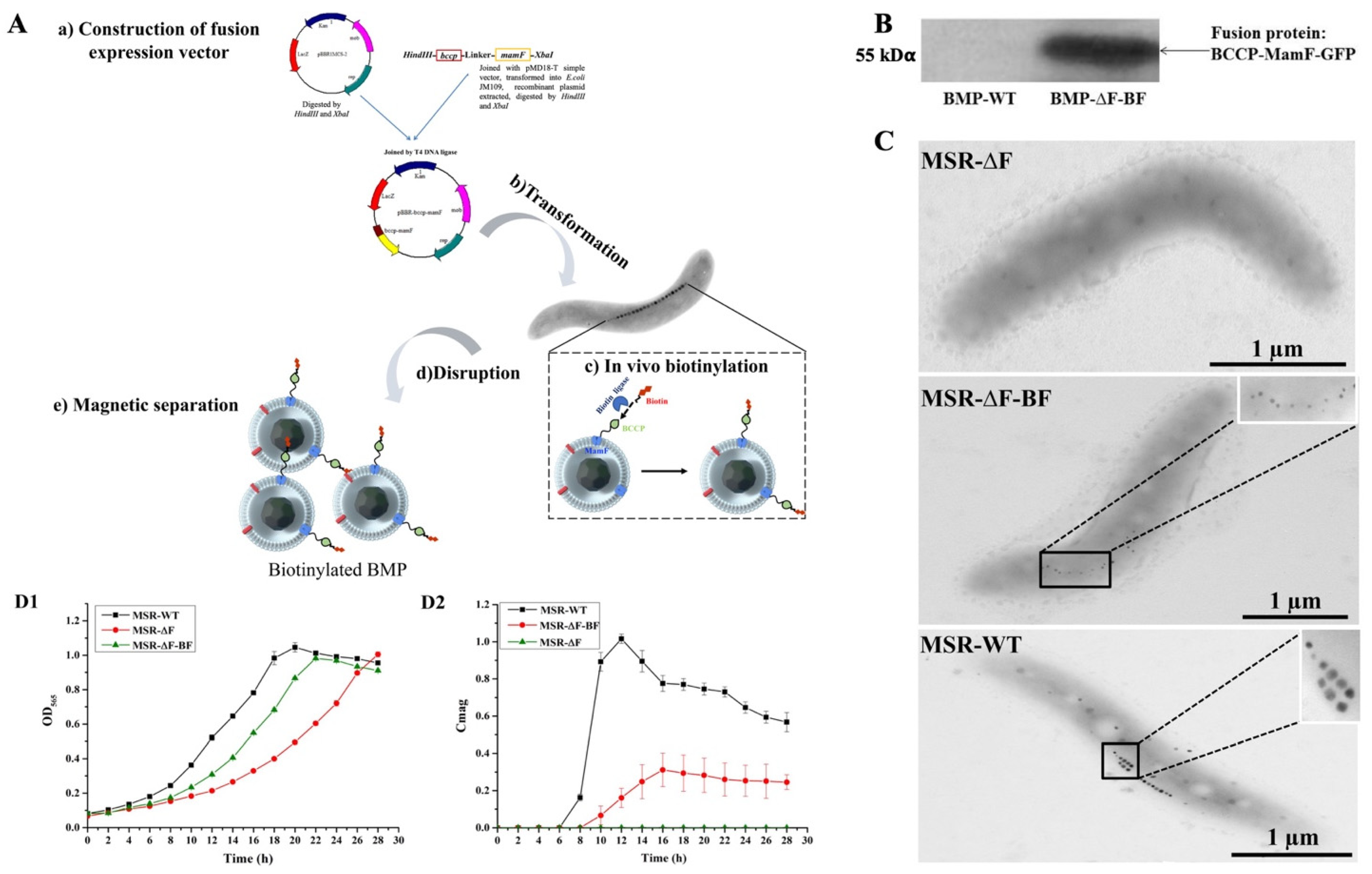
2.2. Construction of Recombinant Strains MSR−∆F−BF
2.3. Transmission Electron Microscope (TEM) Observation
2.4. Submerged Culture of M. gryphiswaldense MSR−∆F−BF
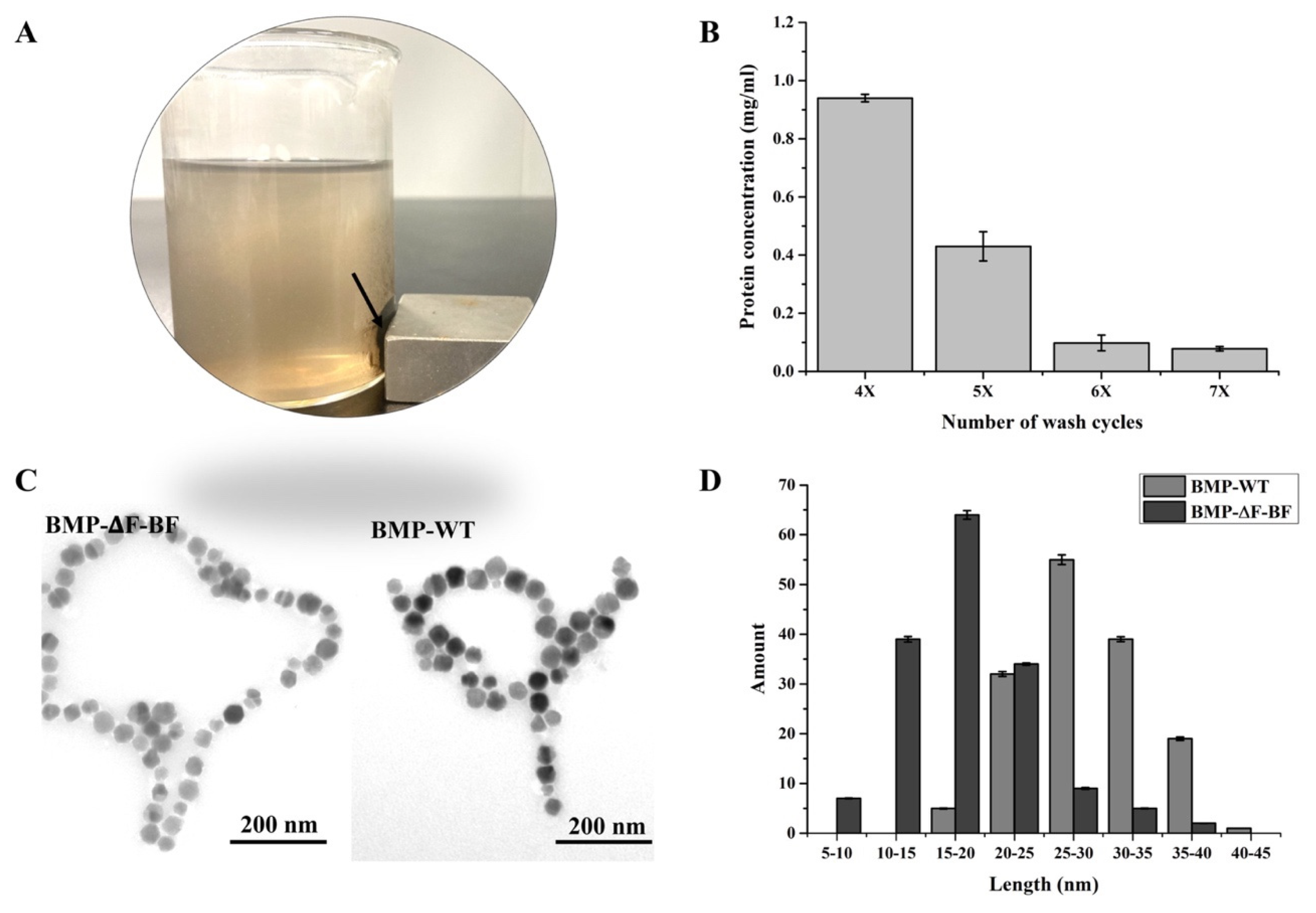
2.5. Extraction and Characterization of BMP−∆F−BF
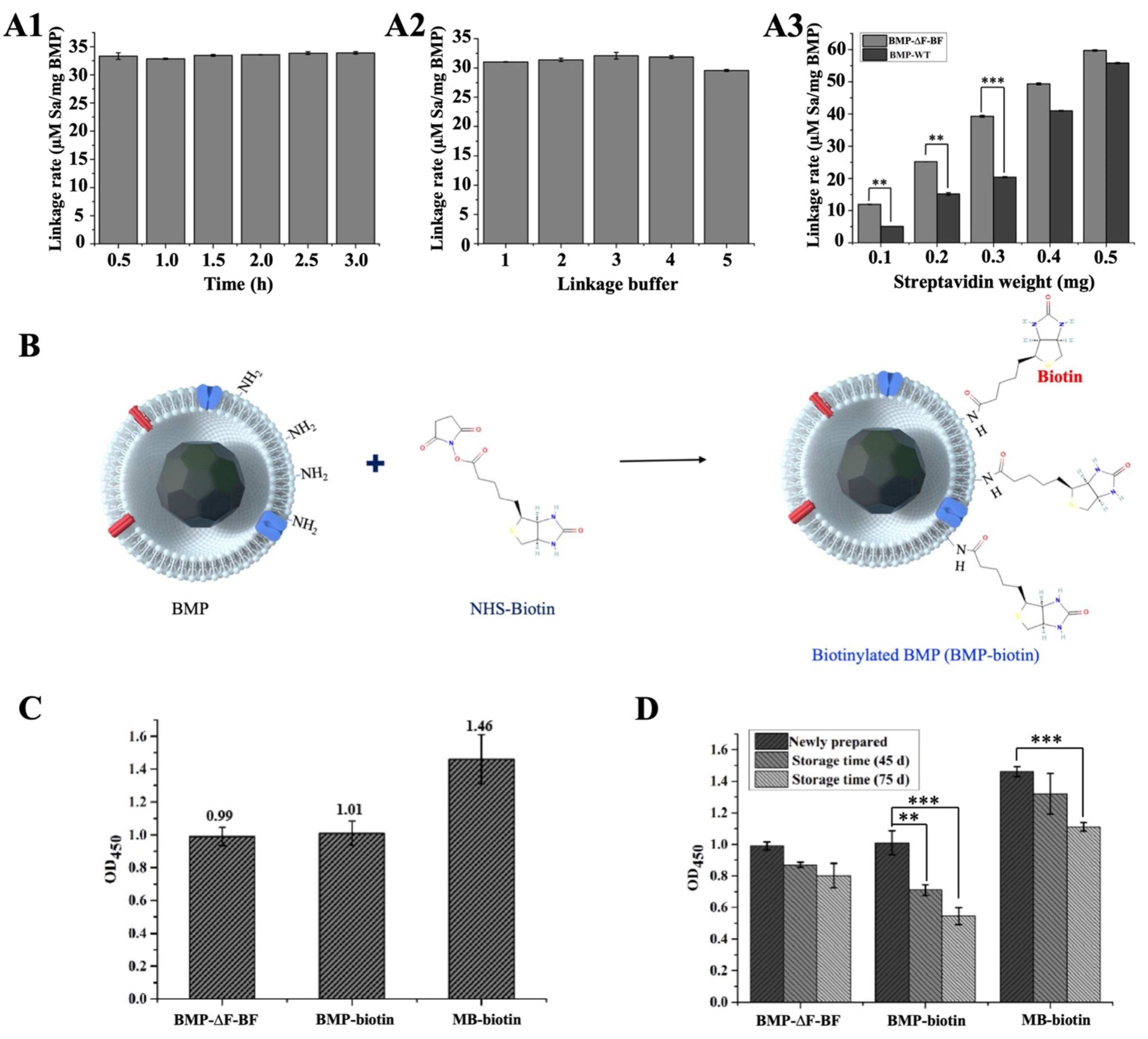
2.6. Preparation of Biotin−Labeled BMPs with NHS−Biotin
2.7. Detection of the Streptavidin−HRP Linkage Rate of BMP−∆F−BF and BMP−biotin
3. Results
3.1. Construction of the Engineered Strain MSR−∆F−BF
3.2. Magnetic Properties Analysis of MSR−∆F−BF
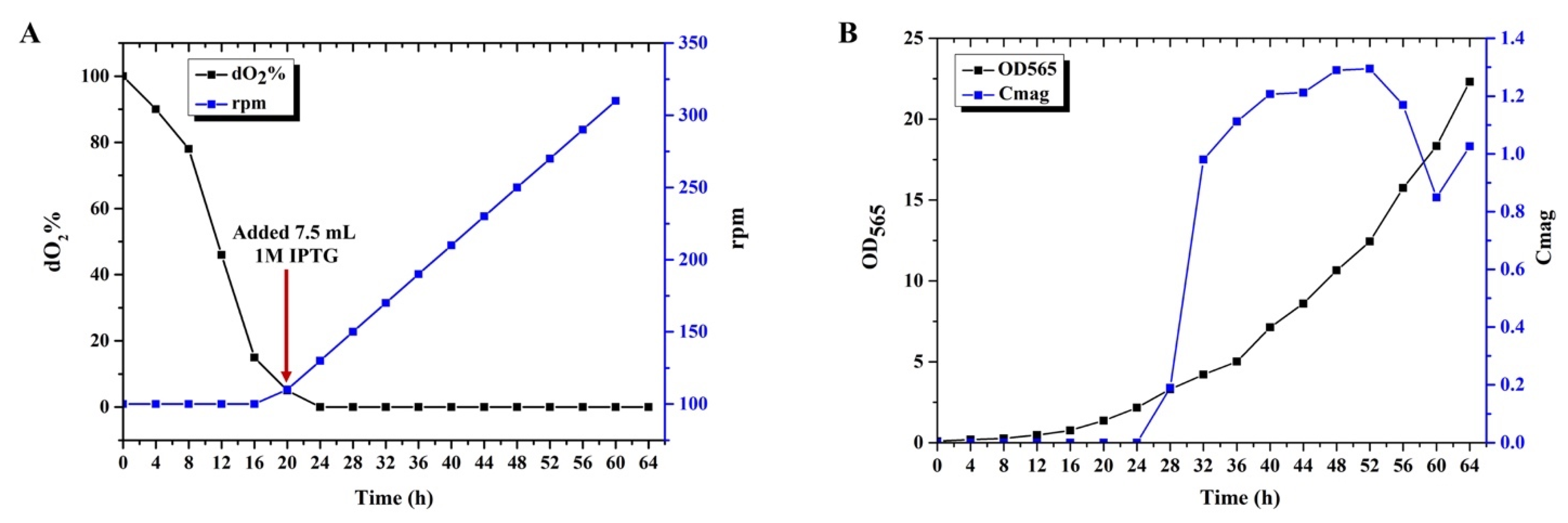
3.3. Submerged Culture of MSR−∆F−BF in Bioreactor
3.4. Preparation and Characterization of BMP−∆F−BF
3.5. Detection of BMP−∆F−BF Coupling Streptavidin Rate
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Clarke, L.; Kitney, R. Developing synthetic biology for industrial biotechnology applications. Biochem. Soc. Trans. 2020, 48, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Karpagam, R.; Jawaharraj, K.; Gnanam, R. Review on integrated biofuel production from microalgal biomass through the outset of transesterification route: A cascade approach for sustainable bioenergy. Sci. Total Environ. 2021, 766, 144236. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Yang, L.; Wang, Z.; Lv, P.; Fu, J.; Miao, C.; Li, M.; Liu, T.; Luo, W. Improved methanol tolerance of Rhizomucor miehei lipase based on Nglycosylation within the alpha-helix region and its application in biodiesel production. Biotechnol. Biofuels 2021, 14, 237. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Atiyeh, H.K.; Li, M.; Chen, Y. Biochar facilitated bioprocessing and biorefinery for productions of biofuel and chemicals: A review. Bioresour. Technol. 2020, 295, 122252. [Google Scholar] [CrossRef] [PubMed]
- Chau, T.H.T.; Nguyen, A.D.; Lee, E.Y. Boosting the acetol production in methanotrophic biocatalyst Methylomonas sp. DH-1 by the coupling activity of heteroexpressed novel protein PmoD with endogenous particulate methane monooxygenase. Biotechnol. Biofuels Bioprod. 2022, 15, 7. [Google Scholar] [CrossRef] [PubMed]
- Andrews, F.; Faulkner, M.; Toogood, H.S.; Scrutton, N.S. Combinatorial use of environmental stresses and genetic engineering to increase ethanol titres in cyanobacteria. Biotechnol. Biofuels 2021, 14, 240. [Google Scholar] [CrossRef]
- Sun, Y.; Li, X.; Wu, L.; Li, Y.; Li, F.; Xiu, Z.; Tong, Y. The advanced performance of microbial consortium for simultaneous utilization of glucose and xylose to produce lactic acid directly from dilute sulfuric acid pretreated corn stover. Biotechnol. Biofuels 2021, 14, 233. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Lin, C.J.; Lee, W.C.; Teng, H.Y.; Chuang, M.H. Production of succinic acid through the fermentation of Actinobacillus succinogenes on the hydrolysate of Napier grass. Biotechnol. Biofuels Bioprod. 2022, 15, 9. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Ying, W.; Li, X.; Zhu, J.; Xu, Y.; Zhang, J. Optimized production of xylooligosaccharides from poplar: A biorefinery strategy with sequential acetic acid/sodium acetate hydrolysis followed by xylanase hydrolysis. Bioresour. Technol. 2022, 347, 126683. [Google Scholar] [CrossRef] [PubMed]
- Tay, Z.W.; Chandrasekharan, P.; Chiu-Lam, A.; Hensley, D.W.; Dhavalikar, R.; Zhou, X.Y.; Yu, E.Y.; Goodwill, P.W.; Zheng, B.; Rinaldi, C.; et al. Magnetic Particle Imaging-Guided Heating in vivo Using Gradient Fields for Arbitrary Localization of Magnetic Hyperthermia Therapy. ACS Nano 2018, 12, 3699–3713. [Google Scholar] [CrossRef]
- Wang, X.; Chen, L.; Ge, J.; Afshari, M.J.; Yang, L.; Miao, Q.; Duan, R.; Cui, J.; Liu, C.; Zeng, J.; et al. Rational Constructed Ultra-Small Iron Oxide Nanoprobes Manifesting High Performance for T1-Weighted Magnetic Resonance Imaging of Glioblastoma. Nanomaterials 2021, 11, 2601. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Bai, X.; Jia, L.; Sun, H.; Feng, Y.; Wang, L.; Zhang, C.; Chen, Y.; Ji, Y.; Zhang, D.; et al. Precise Control of Customized Macrophage Cell Robot for Targeted Therapy of Solid Tumors with Minimal Invasion. Small 2021, 17, e2103986. [Google Scholar] [CrossRef] [PubMed]
- Kurt, H.; Pishva, P.; Pehlivan, Z.S.; Arsoy, E.G.; Saleem, Q.; Bayazit, M.K.; Yuce, M. Nanoplasmonic biosensors: Theory, structure, design, and review of recent applications. Anal. Chim. Acta 2021, 1185, 338842. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zhu, X.; Yu, Y.; Li, C.; Zhang, Z.; Wang, F. Nanotechnology Strategies for Plant Genetic Engineering. Adv. Mater. 2021, 34, e2106945. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.J.B.; Bouwmeester, H.; Gottardo, S.; Amenta, V.; Arena, M.; Brandhoff, P.; Marvin, H.J.P.; Mech, A.; Moniz, F.B.; Pesudo, L.Q.; et al. Nanomaterials for products and application in agriculture, feed and food. Trends Food Sci. Technol. 2016, 54, 155–164. [Google Scholar] [CrossRef]
- Finglas, P.M.; Yada, R.Y.; Toldrá, F. Nanotechnology in foods: Science behind and future perspectives. Trends Food Sci. Technol. 2014, 40, 125–126. [Google Scholar] [CrossRef]
- Song, C.; Sun, W.; Xiao, Y.; Shi, X. Ultrasmall iron oxide nanoparticles: Synthesis, surface modification, assembly, and biomedical applications. Drug Discov. Today 2019, 24, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Nabiyouni, G.; Julaee, M.; Ghanbari, D.; Aliabadi, P.C.; Safaie, N. Room temperature synthesis and magnetic property studies of Fe3O4 nanoparticles prepared by a simple precipitation method. J. Ind. Eng. Chem. 2015, 21, 599–603. [Google Scholar] [CrossRef]
- Ahmadi, S.; Fazilati, M.; Nazem, H.; Mousavi, S.M. Green Synthesis of Magnetic Nanoparticles Using Satureja hortensis Essential Oil toward Superior Antibacterial/Fungal and Anticancer Performance. BioMed Res. Int. 2021, 2021, 8822645. [Google Scholar] [CrossRef] [PubMed]
- Kamath, V.; Chandra, P.; Jeppu, G.P. Comparative study of using five different leaf extracts in the green synthesis of iron oxide nanoparticles for removal of arsenic from water. Int. J. Phytoremediation 2020, 22, 1278–1294. [Google Scholar] [CrossRef] [PubMed]
- Arjaghi, S.K.; Alasl, M.K.; Sajjadi, N.; Fataei, E.; Rajaei, G.E. Green Synthesis of Iron Oxide Nanoparticles by RS Lichen Extract and its Application in Removing Heavy Metals of Lead and Cadmium. Biol. Trace Elem. Res. 2021, 199, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Soni, V.; Raizada, P.; Singh, P.; Cuong, H.N.; Saini, A.; Saini, R.V.; Nadda, A.K.; Nguyen, V.H. Sustainable and green trends in using plant extracts for the synthesis of biogenic metal nanoparticles toward environmental and pharmaceutical advances: A review. Environ. Res. 2021, 202, 111622. [Google Scholar] [CrossRef] [PubMed]
- Kezic, S.; Nunez, R.; Hallmann, S.; Havmose, M.S.; Johansen, J.D.; John, S.M.; Macan, M.; Symanzik, C.; Uter, W.; Weinert, P.; et al. Occupational exposure of hairdressers to airborne hazardous chemicals: A scoping review. Int. J. Eenviron. Res. Public Health 2022, 19, 4176. [Google Scholar] [CrossRef] [PubMed]
- Uebe, R.; Schuler, D. Magnetosome biogenesis in magnetotactic bacteria. Nat. Rev. Microbiol. 2016, 14, 621–637. [Google Scholar] [CrossRef]
- Siponen, M.I.; Legrand, P.; Widdrat, M.; Jones, S.R.; Zhang, W.J.; Chang, M.C.; Faivre, D.; Arnoux, P.; Pignol, D. Structural insight into magnetochrome-mediated magnetite biomineralization. Nature 2013, 502, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Lower, B.H.; Bazylinski, D.A. The bacterial magnetosome: A unique prokaryotic organelle. J. Mol. Microbiol. Biotechnol. 2013, 23, 63–80. [Google Scholar] [CrossRef] [PubMed]
- Vargas, G.; Cypriano, J.; Correa, T.; Leao, P.; Bazylinski, D.A.; Abreu, F. Applications of Magnetotactic Bacteria, Magnetosomes and Magnetosome Crystals in Biotechnology and Nanotechnology: Mini-Review. Molecules 2018, 23, 2438. [Google Scholar] [CrossRef] [PubMed]
- Arakaki, A.; Nakazawa, H.; Nemoto, M.; Mori, T.; Matsunaga, T. Formation of magnetite by bacteria and its application. J. R. Soc. Interface 2008, 5, 977–999. [Google Scholar] [CrossRef] [PubMed]
- Basit, A.; Wang, J.; Guo, F.; Niu, W.; Jiang, W. Improved methods for mass production of magnetosomes and applications: A review. Microb. Cell Factories 2020, 19, 197. [Google Scholar] [CrossRef] [PubMed]
- Edouard, A.; Darine, A.H.; Olivier, S.; Maxime, T.; Alain, T.; Nadege, B.; Florence, G.; Franceois, G.; Imene, C. Nanaoprobe synthesized by Magnetotactic bacteria, detecting flourescence variations under dissociation of rhodamine B from magnetosomes following temperature, pH changes, or the application of radiation. ACS Appl. Mater. Interfaces 2017, 9, 36562–36572. [Google Scholar]
- Nan, X.; Teng, Y.; Tian, J.; Hu, Z.; Fang, Q. A comprehensive assessment of the biocompatibility of Magnetospirillum gryphiswaldense MSR-1 bacterial magnetosomes in vitro and in vivo. Toxicology 2021, 462, 152949. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.B.; Tang, T.; Duan, J.H.; Xu, P.X.; Wang, Z.L.; Zhang, Y.; Wu, L.Y.; Li, Y. Biocompatibility of bacterial magnetosome: Acute toxicity, immunotoxicity and cytotoxicity. Nanotoxicology 2020, 4, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Yue, X.X.; Zhang, S.; Chen, P.; Xu, Z.L.; Li, Y.; Li, H.Y. Biocompatibility evaluation of magnetosomes formed by Acidithiobacillus ferrooxidans. Mater. Sci. Eng. C 2012, 32, 1802–1807. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.T.; Liu, J.; Tong, J.Q.; Tang, T.; Kong, W.C.; Wang, X.W.; Li, Y.; Tang, J.T. Heating effect and biocompatibility of bacterial magnetosomes as potential materials used in magnetic fluid hyperthermia. Prog. Nat. Sci. Mater. Int. 2012, 22, 31–39. [Google Scholar] [CrossRef]
- Nan, X.H.; Lai, W.J.; Li, D.; Tian, J.S.; Hu, Z.Y.; Fang, Q.J. Biocompatibility of bacterial magnetosomes as MRI contrast agent: A Long-Term in vivo Follow-Up study. Nanomaterials 2021, 11, 1235. [Google Scholar] [CrossRef] [PubMed]
- Boucher, M.; Geffroy, F.; Bellanger, L.; Selingue, E.; Pignol, D.; Ginet, N.; Meriaux, S. Genetically tailored magnetosomes used as MRI probe for molecular imaging of brain tumor. Biomaterials 2017, 121, 167–178. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Tian, J.; Xu, J.; Wang, K.; Li, J.; Gee, S.J.; Hammock, B.D.; Li, Q.X.; Xu, T. Strong and oriented conjugation of nanobodies onto magnetosomes for the development of a rapid immunomagnetic assay for the environmental detection of tetrabromobisphenol-A. Anal. Bioanal. Chem. 2018, 410, 6633–6642. [Google Scholar] [CrossRef]
- Xiang, Z.; Yang, X.; Xu, J.; Lai, W.; Wang, Z.; Hu, Z.; Tian, J.; Geng, L.; Fang, Q. Tumor detection using magnetosome nanoparticles functionalized with a newly screened EGFR/HER2 targeting peptide. Biomaterials 2017, 115, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Geng, Y.; Zhang, Y.; Wang, X.; Liu, J.; Basit, A.; Miao, T.; Liu, W.; Jiang, W. Bacterial magnetosomes loaded with doxorubicin and transferrin improve targeted therapy of hepatocellular carcinoma. Nanotheranostics 2019, 3, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Hu, J.; Liu, L.; Li, L.; Wang, X.; Zhang, H.; Jiang, W.; Tian, J.; Li, Y.; Li, J. Surface expression of protein A on magnetosomes and capture of pathogenic bacteria by magnetosome/antibody complexes. Front. Microbiol. 2014, 5, 136. [Google Scholar] [CrossRef] [PubMed]
- Mickoleit, F.; Lanzloth, C.; Schuler, D. A Versatile Toolkit for Controllable and Highly Selective Multifunctionalization of Bacterial Magnetic Nanoparticles. Small 2020, 16, e1906922. [Google Scholar] [CrossRef] [PubMed]
- Honda, T.; Tanaka, T.; Yoshino, T. Stoichiometrically Controlled Immobilization of Multiple Enzymes on Magnetic Nanoparticles by the Magnetosome Display System for Efficient Cellulose Hydrolysis. Biomacromolecules 2015, 16, 3863–3868. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ni, Q.; Xu, C.; Wan, B.; Gang, Y.; Zheng, G.; Yang, Z.; Tao, J.; Wen, J.; Zhang, J.; et al. Smart bacterial magnetic nanoparticles for Tumor-Targeting magnetic resonance imaging of HER2-postive breast cancer. ACS Appl. Mater. Interfaces 2019, 11, 2654–3665. [Google Scholar]
- Xu, J.; Liu, L.; He, J.; Ma, S.; Li, S.; Wang, Z.; Xu, T.; Jiang, W.; Wen, Y.; Li, Y.; et al. Engineered magnetosomes fused to functional molecule (protein A) provide a highly effective alternative to commercial immunomagnetic beads. J. Nanobiotechnology 2019, 17, 37. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Takeyama, H.; Tanaka, T.; Matsunaga, T. Effects of growth medium composition, iron sources and atmospheric oxygen concentrations on production of luciferase-bacterial magnetic particle comples by a recombinant Magnetospirillum magneticum AMB-1. Enzym. Microb. Technol. 2001, 29, 13–19. [Google Scholar] [CrossRef]
- Fernandez-Suarez, M.; Chen, T.S.; Ting, A.Y. Protein-protein interaction detection in vitro and in cells by proximity biotinylation. J. Am. Chem. Soc. 2008, 130, 9251–9253. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.; Stern, D.; Seeber, F. Expression of in vivo biotinylated recombinant antigens SAG1 and SAG2A from Toxoplasma gondii for improved seroepidemiological bead-based multiplex assays. BMC Biotechnol. 2020, 20, 53. [Google Scholar] [CrossRef] [PubMed]
- Boer, E.; Rodriguez, P.; Bonte, E.; Krijgsveld, J.; Katsantoni, E.; Heck, A.; Grosveld, F.; Strouboulis, J. Efficient biotinylation and single-step purification of tagged transcription factors in mammalian cells and transgenic mice. Proc. Natl. Acad. Sci. USA 2003, 100, 7480–7485. [Google Scholar] [CrossRef]
- Maeda, Y.; Yoshino, T.; Matsunaga, T. Novel nanocomposites consisting of in vivo-biotinylated bacterial magnetic particles and quantum dots for magnetic separation and fluorescent labeling of cancer cells. J. Mater. Chem. 2009, 19, 6361–6366. [Google Scholar] [CrossRef]
- Zwiener, T.; Dziuba, M.; Mickoleit, F.; Ruckert, C.; Busche, T.; Kalinowski, J.; Uebe, R.; Schuler, D. Towards a ‘chassis’ for bacterial magnetosome biosynthesis: Genome streamlining of Magnetospirillum gryphiswaldense by multiple deletions. Microb. Cell Factories 2021, 20, 35. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Simon, R.; Priefer, U.; Puhler, A. A broad host range mobilization system for in vivo genetic engineering: Transposon mutagenesis in Gram negative bacteria. Bio/Technol. 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Ali, I.; Peng, C.; Khan, Z.M.; Naz, I. Yield cultivation of magnetotactic bacteria and magnetosomes: A review. J. Basic Microbiol. 2017, 57, 643–652. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Jiang, W.; Li, Y.; Li, J. Semicontinuous Culture of Magnetospirillum gryphiswaldense MSR-1 Cells in an Autofermentor by Nutrient-Balanced and Isosmotic Feeding Strategies. Appl. Environ. Microbiol. 2011, 77, 5851–5856. [Google Scholar] [CrossRef] [PubMed]
- Joudeh, N.; Linke, D. Nanoparticle classification, physicochemical properties, characterization, and applications: A comprehensive review for biologists. J. Nanobiotechnology 2022, 20, 262. [Google Scholar] [CrossRef] [PubMed]
- Park, M. Surface Display Technology for Biosensor Applications: A Review. Sensors 2020, 20, 2775. [Google Scholar] [CrossRef] [PubMed]
- Kunamneni, A.; Ogaugwu, C.; Bradfute, S.; Durvasula, R. Ribosome Display Technology: Applications in Disease Diagnosis and Control. Antibodies 2020, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Kamalinia, G.; Grindel, B.J.; Takahashi, T.T.; Millward, S.W.; Roberts, R.W. Directing evolution of novel ligands by mRNA display. Chem. Soc. Rev. 2021, 50, 9055–9103. [Google Scholar] [CrossRef]
- Chaiet, L.; Wolf, F.J. The properties of streptavidin, a biotin binding protein produced by Streptomycetes. Arch. Biochem. Biophys. 1964, 106, 1–5. [Google Scholar] [CrossRef]
- Lombard, J.; Moreira, D. Early evolution of the biotin dependent carboxylase family. BMC Evol. Biol. 2011, 11, 232. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Elizondo-Noriega, A.; Cantu, D.C.; Reilly, P.J. Structural classification of biotin carboxyl carrier proteins. Biotechnol. Lett. 2012, 34, 1869–1875. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Cheng, K. The Principles and Applications of Avidin-Based Nanoparticles in Drug Delivery and Diagnosis. J. Control. Release 2017, 245, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Han, J.; Uhm, S.; Jang, Y.; Kang, C.; Kim, J.H.; Kim, J. Recent Development of Biotin Conjugation in Biological Imaging, Sensing, and Target Delivery. Chem. Commun. 2015, 51, 10403–10418. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.F.; Wu, P.; Jiang, Z.H.; Wei, X.Y. Synthesis and Tumor Cell Growth Inhibitory Activity of Biotinylated Annonaceous Acetogenins. Eur. J. Med. Chem. 2014, 71, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhao, X.; Chen, J.; Chen, J.; Kuznetsova, L.; Wong, S.S.; Ojima, I. Mechanism-Based Tumor-Targeting Drug Delivery System. Validation of Efficient Vitamin Receptor-Mediated Endocytosis and Drug Release. Bioconjugate Chem. 2010, 21, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Khramtsov, P.; Kropaneva, M.; Bochkova, M.; Kiselkov, D.; Timganova, V.; Zamorina, S.; Rayev, M. Nuclear magnetic resonance immunoassay of tetanus antibodies based on the displacement of magnetic nanoparticles. Anal. Bioanal. Chem. 2021, 413, 1461–1471. [Google Scholar] [CrossRef] [PubMed]
- Nosrati, H.; Barzegari, P.; Danafar, H.; Manjili, H.K. Biotin-Functionalized Copolymeric PEG-PCL Micelles for in vivo TumourTargeted Delivery of Artemisinin. Artif. Cells Nanomed. Biotechnol. 2019, 47, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; van Steenbergen, M.J.; Beztsinna, N.; Shi, Y.; Lammers, T.; van Nostrum, C.F.; Hennink, W.E. Biotin-Decorated All-HPMA Polymeric Micelles for Paclitaxel Delivery. J. Control. Release 2020, 328, 970–984. [Google Scholar] [CrossRef] [PubMed]
- Rompicharla, S.V.K.; Kumari, P.; Bhatt, H.; Ghosh, B.; Biswas, S. Biotin Functionalized PEGylated Poly (Amidoamine) Dendrimer Conjugate for Active Targeting of Paclitaxel in Cancer. Int. J. Pharm. 2019, 557, 329–341. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain and Plasmid | Description | Source or Reference |
|---|---|---|
| Strains | ||
| Magnetospirillum gryphiswaldense MSR-1 | Wild-type magnetotactic bacteria, Nxr | DSM 6361 |
| Magnetospirillum gryphiswaldense MSR−∆F | mamF-defective mutant, Nxr Gmr | Ref [40] |
| Magnetospirillum gryphiswaldense MSR−∆F−BF | mamF mutant harboring pBBR−bccp−mamF, Nxr Gmr Kmr | This study |
| E. coli DH5α | endA1 hsdR17(rK− mK+) supE44 thi-1 recA1 gyrA (Nalr) recA1 ∆(lacZYA-argF)U169 deoR[Φ80dlacZ∆M15] | Ref [51] |
| E. coli S17-1 | thi endA recA hsdR with RP4-2-Tc::Mu-Km::Tn7 integrated into the chromosome; Smr | Ref [52] |
| Plasmids | ||
| pMD18-T simple | Clone vector, Ampr | Takara |
| pBBR1MCS-2 | Broad-host-range shuttle vector, Kmr | Lab collection |
| pBBR−bccp−mamF-gfp | pBBR1MCS-2 containing bccp-mamF-gfp, Kmr | This study |
| pBBR−bccp−mamF | pBBR1MCS-2 containing bccp-mamF, Kmr | This study |
| Strains | Coercivity (Hc, mT) | Saturation Magnetization (Ms, µ Am2) |
|---|---|---|
| MSR−∆F−BF | 2.24 | 4.26 |
| MSR−WT | 7.65 | 12.55 |
| Magnetosomes/ Magnetic Beads | Hydrated Radii (nm) | Zeta Potentials (mV) | Polydispersity |
|---|---|---|---|
| BMP–WT | 113.9 ± 62.19 | −17.7 ± 6.78 | 0.48 |
| BMP–∆F–BF | 53.52 ± 10.23 | −23.6 ± 6.12 | 0.35 |
| MB–biotin | 669.3 ± 288.5 | −17.5 ± 3.84 | 0.59 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Ma, S.; Zheng, H.; Pang, B.; Li, S.; Li, F.; Feng, L.; Tian, J. Biomanufacturing Biotinylated Magnetic Nanomaterial via Construction and Fermentation of Genetically Engineered Magnetotactic Bacteria. Bioengineering 2022, 9, 356. https://doi.org/10.3390/bioengineering9080356
Xu J, Ma S, Zheng H, Pang B, Li S, Li F, Feng L, Tian J. Biomanufacturing Biotinylated Magnetic Nanomaterial via Construction and Fermentation of Genetically Engineered Magnetotactic Bacteria. Bioengineering. 2022; 9(8):356. https://doi.org/10.3390/bioengineering9080356
Chicago/Turabian StyleXu, Junjie, Shijiao Ma, Haolan Zheng, Bo Pang, Shuli Li, Feng Li, Lin Feng, and Jiesheng Tian. 2022. "Biomanufacturing Biotinylated Magnetic Nanomaterial via Construction and Fermentation of Genetically Engineered Magnetotactic Bacteria" Bioengineering 9, no. 8: 356. https://doi.org/10.3390/bioengineering9080356
APA StyleXu, J., Ma, S., Zheng, H., Pang, B., Li, S., Li, F., Feng, L., & Tian, J. (2022). Biomanufacturing Biotinylated Magnetic Nanomaterial via Construction and Fermentation of Genetically Engineered Magnetotactic Bacteria. Bioengineering, 9(8), 356. https://doi.org/10.3390/bioengineering9080356