
Preliminary Examination of the Toxicity of Spalting Fungal Pigments: A Comparison between Extraction Methods

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Pigment Samples
2.1.1. Solvent-Extracted Pigment
2.1.2. Pigments from Liquid Media
2.1.3. Solid Pigments
2.1.4. Solvent-Extracted Pigment Concentration
2.2. Zebrafish Preparation and Exposure for Pigment Extracts
2.2.1. Pigment from Solvent Extraction
2.2.2. Pigment from Liquid Culture
2.2.3. Assessment Protocol
2.3. Zebrafish Preparation and Exposure for Solid Purified Pigments
2.3.1. Preparation of Zebrafish
2.3.2. Exposure Protocol
2.3.3. Embryo Photomotor Response (EPR) Behavior
2.3.4. Larval Photomotor Response Behavior (LPR)
2.3.5. Mortality and Morphology Response
2.4. Statistical Analysis
2.4.1. Zebrafish Exposure to Pigment Extracts
2.4.2. Zebrafish Exposure to Solid Pigments
3. Results and Discussion
3.1. Solvent-Extracted Pigment and Liquid Culture Testing
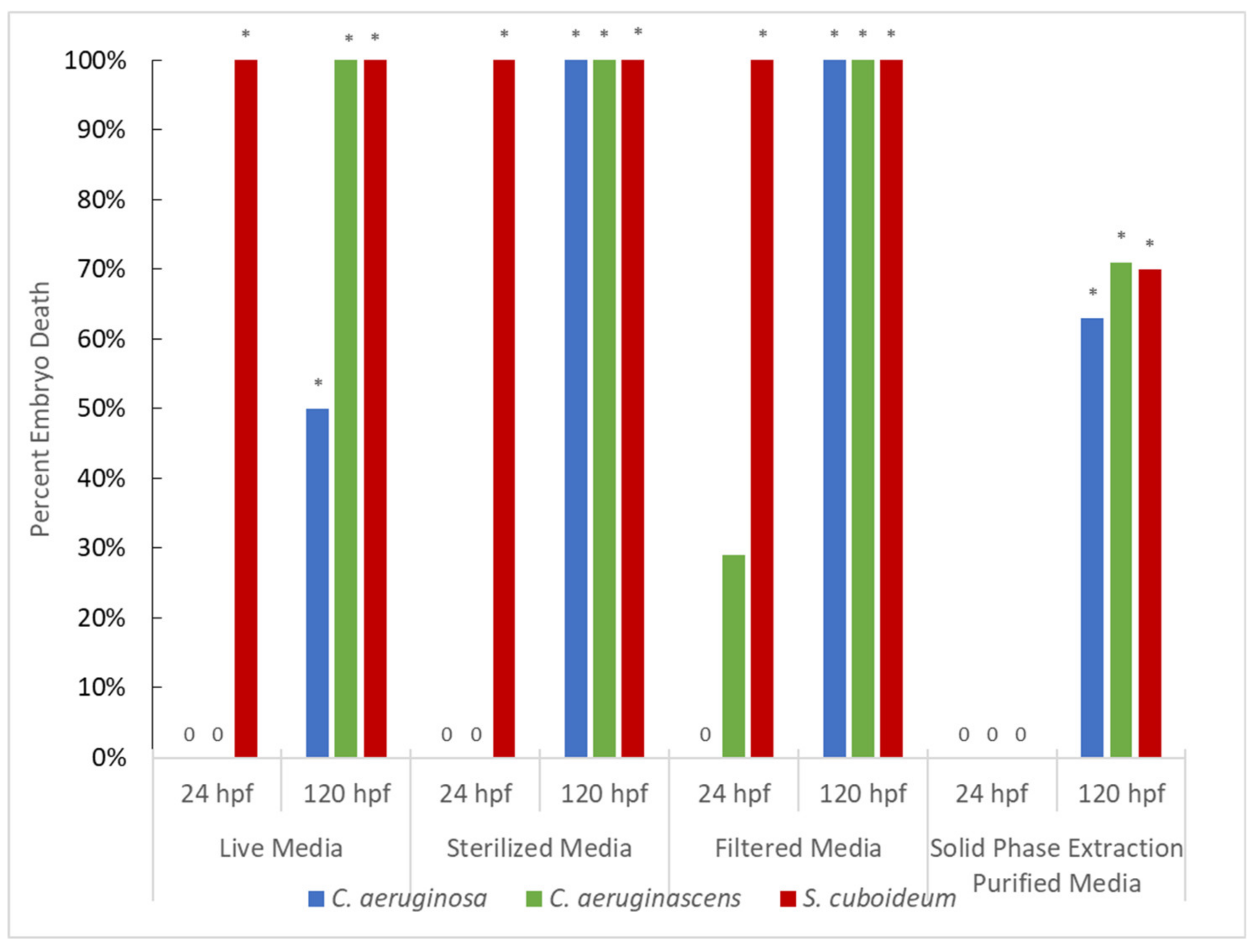
3.1.1. Pigment from Solvent Extraction (No Purification)
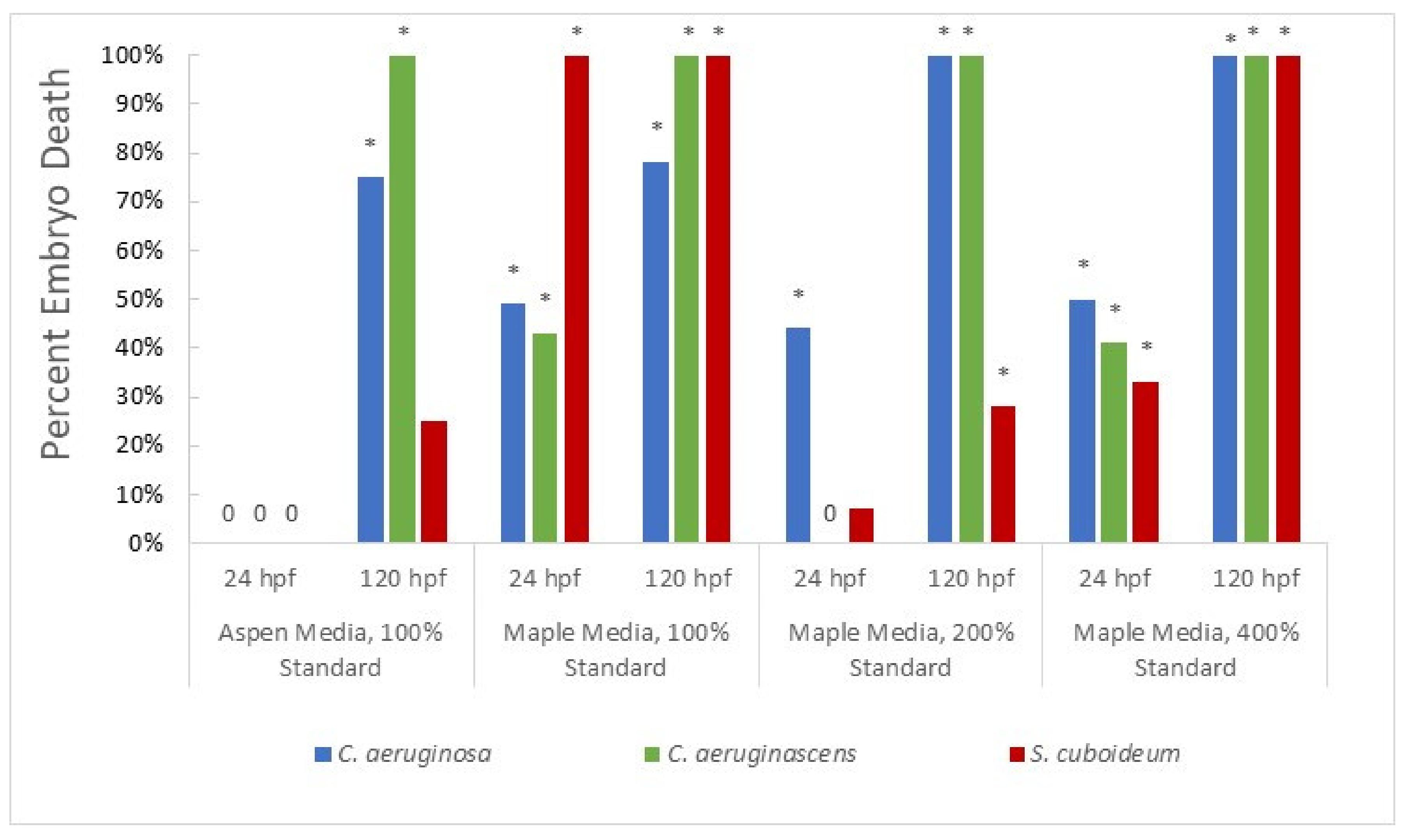
3.1.2. Pigment from Liquid Culture
3.2. Solid Pigments and Behavior Response Testing
3.2.1. Embryo Photomotor Response
3.2.2. Larval Photomotor Response
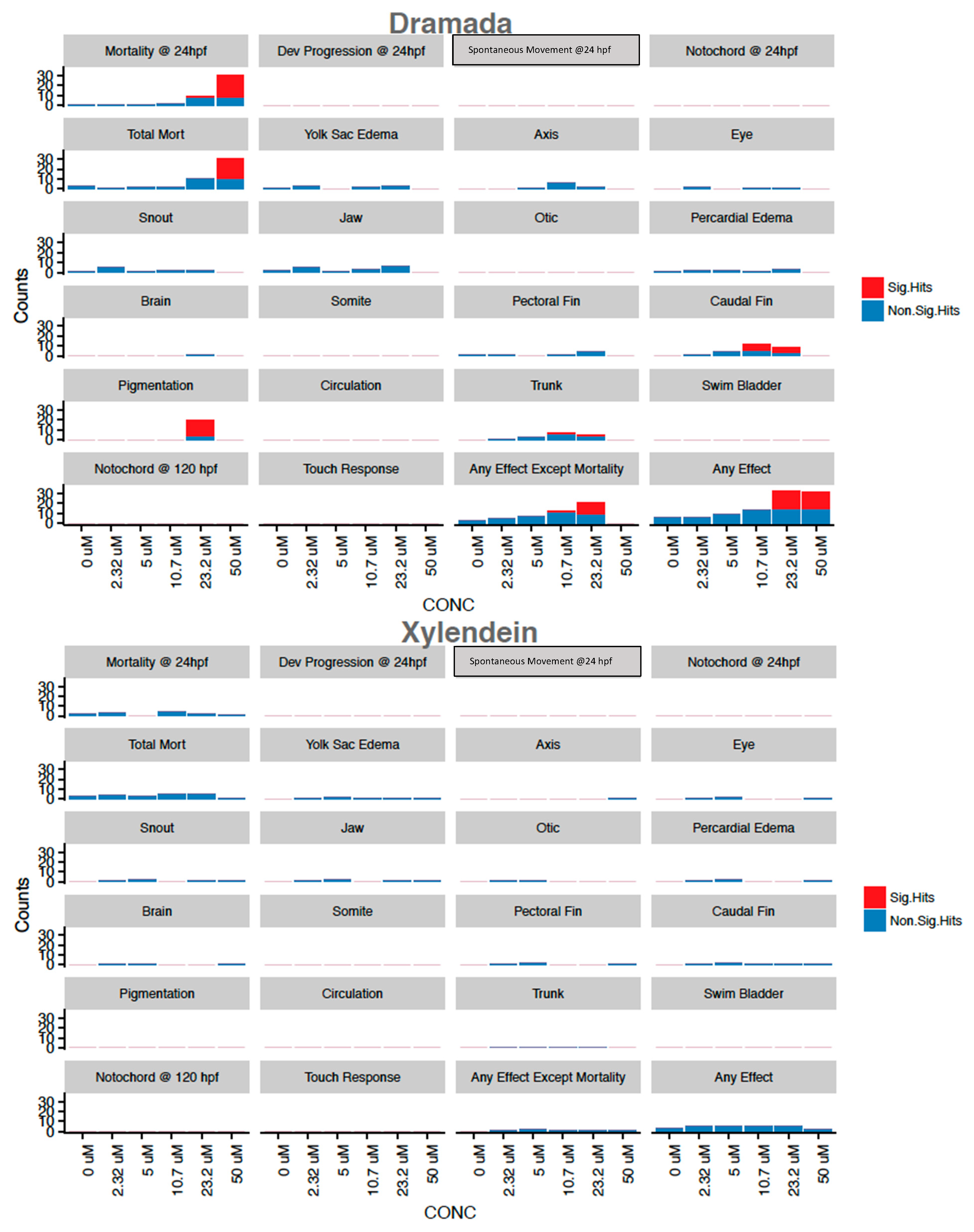
3.2.3. Mortality and Morphology
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robinson, S.C.; Michaelsen, H.; Robinson, J.C. Spalted wood. The History, Science and Art of a Unique Material, 1st ed.; Schiffer Publishing, Ltd.: Atglen, PA, USA, 2016; p. 287. [Google Scholar]
- Seaver, F.G. Photographs and descriptions of cup fungi –XXIV. Chlorociboria. Mycologia 1936, 28, 309–394. [Google Scholar]
- Dixon, J.R. Chlorosplenium and its segregates. The genera chlorociboria and chlorencoelia. Mycotaxon 1975, 1, 193–237. [Google Scholar]
- Oeder, G.C. Flora Danica. Verlegts Heineck, Mumme und Faber; Nicolaus Möller: Kopenhagen, Denmark, 1770. [Google Scholar]
- Chidester, M.S. A pink stain of wood caused by a species of geotrichum. Phytopathology 1940, 30, 530–533. [Google Scholar]
- Kang, H.; Sigler, L.; Lee, J.; Gibas, C.; Yun, S.; Lee, Y. Xylogone ganodermophthora sp. nov., an ascomycetous pathogen causing yellow rot on cultivated mushroom Ganoderma lucidum in Korea. Mycologia 2010, 102, 1167–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchette, R.A.; Wilmering, A.M.; Baumeister, M. The use of green-stained wood caused by the fungus Chlorociboria in Intarsia masterpieces from the 15th century. Holzforschung 1992, 46, 225–232. [Google Scholar] [CrossRef]
- Vega Gutierrez, T.P.; Robinson, C.S. Determining the Presence of Spalted Wood in Spanish Marquetry Woodworks of the 1500s through the 1800s. Coatings 2017, 7, 188. [Google Scholar] [CrossRef] [Green Version]
- Otterstedt, A. Investigating green Marquetry on bowed-string instruments. The leaves be greene. Galpin Soc. J. 2001, 330–338. [Google Scholar] [CrossRef]
- Rommier, P.T.A. Sur un nouvelle matière colorante appelée xylindeine et extraite de certains bois morts. Comptes Rendus Hebd. Des Séances De L’académie Des Sci. 1868, 66, 108–109. [Google Scholar]
- Blackburn, G.M.; Ekong, D.E.; Nielson, A.H.; Todd, L. Xylindein. Chimia 1965, 19, 208–212. [Google Scholar]
- Edwards, R.L.; Kale, N. The structure of xylindein. Tetrahedron 1965, 21, 2095–2107. [Google Scholar] [CrossRef]
- Saikawa, Y.; Watanabe, T.; Hashimoto, K.; Nakata, A. Absolute configuration and tautomeric structure of xylindein, a blue-green pigment of Chlorociboria species. Phytochemistry 2000, 55, 237–240. [Google Scholar] [CrossRef]
- Harrison, R.; Quinn, A.; Weber, G.; Johnson, B.; Rath, J.; Remcho, V.; Robinson, S.; Ostroverkhovaa, O. Fungi-Derived Pigments as Sustainable Organic (Opto)electronic Materials. Available online: https://www.spiedigitallibrary.org/conference-proceedings-of-spie/10101/1/Fungi-derived-pigments-as-sustainable-organic-optoelectronic-materials/10.1117/12.2251265.short?SSO=1 (accessed on 20 February 2021).
- Vega Gutierrez, M.S.; Hazell, K.K.; Simonsen, J.; Robinson, C.S. Description of a Naphthoquinonic Crystal Produced by the Fungus Scytalidium cuboideum. Molecules 2018, 23, 1905. [Google Scholar] [CrossRef] [Green Version]
- Gerber, N.N.; Wieclawek, B. The Structures of Two Naphthoquinone Pigments from an Actinomycete1. J. Org. Chem. 1966, 31, 1496–1498. [Google Scholar] [CrossRef]
- Robinson, S.C.; Hinsch, E.; Weber, G.; Leipus, K.; Cerney, D. Wood Colorization through Pressure Treating: The Potential of Extracted Colorants from Spalting Fungi as a Replacement for Woodworkers’ Aniline Dyes. Materials 2014, 7, 5427–5437. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.C.; Vega Gutierrez, S.M.; Garcia, R.A.C.; Iroume, N.; Vorland, N.R.; Andersen, C.; de Oliveira Xaxa, I.D.; Kramer, O.E.; Huber, M.E. Potential for fungal dyes as colorants in oil and acrylic paints. J. Coat. Technol. Res. 2018, 15, 845–849. [Google Scholar] [CrossRef]
- Hinsch, E.M. A Comparative Analysis of Extracted Fungal Pigments and Commercially Available Dyes for Colorizing Textiles. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 2015. [Google Scholar]
- Hinsch, E.; Robinson, S. Comparing Colorfastness to Light of Wood-Staining Fungal Pigments and Commercial Dyes: An Alternative Light Test Method for Color Fastness. Coatings 2018, 8, 189. [Google Scholar] [CrossRef] [Green Version]
- Weber, G.; Chen, H.-L.; Hinsch, E.; Freitas, S.; Robinson, S. Pigments extracted from the wood-staining fungi Chlorociboria aeruginosa, Scytalidium cuboideum, and S. ganodermophthorum show potential for use as textile dyes. Coloration Technol. 2014, 130, 445–452. [Google Scholar] [CrossRef]
- Khan, S.; Malik, A. Environmental and Health Effects of Textile Industry Wastewater. In Environmental Deterioration and Human Health: Natural and anthropogenic determinants; Malik, A., Grohmann, E., Akhtar, R., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2014; pp. 55–71. [Google Scholar] [CrossRef]
- Markandeya, S.P.; Shukla, S.P.; Mohan, D. Toxicity of Disperse Dyes and its Removal from Wastewater Using Various Adsorbents: A Review. Res. J. Environ. Toxicol. 2017, 11, 72–89. [Google Scholar]
- Sharma, K.P.; Sharma, S.; Sharma, S.; Singh, P.K.; Kumar, S.; Grover, R.; Sharma, P.K. A comparative study on characterization of textile wastewaters (untreated and treated) toxicity by chemical and biological tests. Chemosphere 2007, 69, 48–54. [Google Scholar] [CrossRef]
- Khan, R.; Bhawana, P.; Fulekar, M.H. Microbial decolorization and degradation of synthetic dyes: A review. Rev. Environ. Sci. Bio/Technol. 2013, 12, 75–97. [Google Scholar] [CrossRef]
- Giesbers, G.; Van Schenck, J.; Vega Gutierrez, M.S.; Robinson, S.; Ostroverkhovaa, O. Fungi-Derived Pigments for Sustainable Organic (Opto)Electronics. Mrs Adv. 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Radić, N.; Štrukelj, B. Endophytic fungi—The treasure chest of antibacterial substances. Phytomedicine 2012, 19, 1270–1284. [Google Scholar] [CrossRef] [PubMed]
- Beekman, A.M.; Barrow, R.A. Fungal Metabolites as Pharmaceuticals. Aust. J. Chem. 2014, 67, 827–843. [Google Scholar] [CrossRef]
- Moss, M.O. Mycotoxins of Aspergillus and other filamentous fungi. J. Appl. Bacteriol. 1989, 67, 69s–81s. [Google Scholar] [CrossRef]
- Narsing Rao, M.P.; Xiao, M.; Li, W.-J. Fungal and Bacterial Pigments: Secondary Metabolites with Wide Applications. Front. Microbiol. 2017, 8, 1113. [Google Scholar] [CrossRef]
- Dufossé, L.; Fouillaud, M.; Caro, Y.; Mapari, S.A.S.; Sutthiwong, N. Filamentous fungi are large-scale producers of pigments and colorants for the food industry. Curr. Opin. Biotechnol. 2014, 26, 56–61. [Google Scholar] [CrossRef]
- Dubravka, F.; Maja, P. Toxicological Properties of Citrinin. Arch. Ind. Hyg. Toxicol. 2009, 60, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, J.C.d.; Oishi, B.O.; Pandey, A.; Soccol, C.R. Biopigments from Monascus: Strains selection, citrinin production and color stability. Braz. Arch. Biol. Technol. 2005, 48, 885–894. [Google Scholar] [CrossRef] [Green Version]
- Liang, B.; Du, X.-J.; Li, P.; Sun, C.-C.; Wang, S. Investigation of Citrinin and Pigment Biosynthesis Mechanisms in Monascus purpureus by Transcriptomic Analysis. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Marič, A.; Skočaj, M.; Likar, M.; Sepčić, K.; Cigić, I.K.; Grundner, M.; Gregori, A. Comparison of lovastatin, citrinin and pigment production of different Monascus purpureus strains grown on rice and millet. J. Food Sci. Technol. 2019, 56, 3364–3373. [Google Scholar] [CrossRef] [PubMed]
- Panda, B.P.; Ali, M. Reduction of citrinin biosynthesis by fatty acids in Monascus fermented food. World Mycotoxin J. 2012, 5, 163–167. [Google Scholar] [CrossRef]
- Zhen, Z.; Xiong, X.; Liu, Y.; Zhang, J.; Wang, S.; Li, L.; Gao, M. NaCl Inhibits Citrinin and Stimulates Monascus Pigments and Monacolin K Production. Toxins (Basel) 2019, 11, 118. [Google Scholar] [CrossRef] [Green Version]
- Gibson, S. A Warning on Spalted Wood. Fine Woodworking 1995, 118, 110–111. [Google Scholar]
- Robinson, S.C. Spalted Wood: Health and Safety. Am. Woodturn. 2011. Available online: https://www.google.com.hk/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&ved=2ahUKEwjMmK_YpPzuAhVCQKwKHUbJAJ0QFjAAegQIBxAD&url=https%3A%2F%2Fwww.peaceriverwoodturners.org%2Fresources%2FDocuments%2FUNIQUE%2520TECHNIQUES%2FSpalted%2520Wood%2520Safety.pdf&usg=AOvVaw0XAz6Tj2Ow1izwSMtkPQmL (accessed on 21 February 2021).
- Ali, S.; Champagne, D.L.; Spaink, H.P.; Richardson, M.K. Zebrafish embryos and larvae: A new generation of disease models and drug screens. Birth Defects Res. Part C: Embryo Today: Rev. 2011, 93, 115–133. [Google Scholar] [CrossRef]
- Kari, G.; Rodeck, U.; Dicker, A.P. Zebrafish: An Emerging Model System for Human Disease and Drug Discovery. Clin. Pharmacol. Ther. 2007, 82, 70–80. [Google Scholar] [CrossRef]
- Hagstrom, D.; Truong, L.; Zhang, S.; Tanguay, R.; Collins, E.-M.S. Comparative Analysis of Zebrafish and Planarian Model Systems for Developmental Neurotoxicity Screens Using an 87-Compound Library. Toxicol. Sci. 2018, 167, 15–25. [Google Scholar] [CrossRef]
- Detrich, H.W.; Westerfield, M.; Zon, L.I. Chapter 1 Overview of the Zebrafish System. In Methods in Cell Biology; Detrich, H.W., Westerfield, M., Zon, L.I., Eds.; Academic Press: Cambridge, MA, USA, 1998; Volume 59, pp. 3–10. [Google Scholar]
- Zon, L.I.; Peterson, R.T. In vivo drug discovery in the zebrafish. Nat. Rev. Drug Discov. 2005, 4, 35–44. [Google Scholar] [CrossRef]
- MacRae, C.A.; Peterson, R.T. Zebrafish as tools for drug discovery. Nat. Rev. Drug Discov. 2015, 14, 721. [Google Scholar] [CrossRef]
- Gehrig, J.; Pandey, G.; Westhoff, J.H. Zebrafish as a Model for Drug Screening in Genetic Kidney Diseases. Front. Pediatrics 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Willett, C.; Fremgen, T. Zebrafish: An Animal Model for Toxicological Studies. Curr. Protoc. Toxicol. 2003, 17, 1.7.1–1.7.18. [Google Scholar] [CrossRef]
- Embry, M.R.; Belanger, S.E.; Braunbeck, T.A.; Galay-Burgos, M.; Halder, M.; Hinton, D.E.; Léonard, M.A.; Lillicrap, A.; Norberg-King, T.; Whale, G. The fish embryo toxicity test as an animal alternative method in hazard and risk assessment and scientific research. Aquat. Toxicol. 2010, 97, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Scholz, S. Zebrafish embryos as an alternative model for screening of drug-induced organ toxicity. Arch. Toxicol. 2013, 87, 767–769. [Google Scholar] [CrossRef] [Green Version]
- Challal, S.; Bohni, N.; Buenafe, O.E.; Esguerra, C.V.; de Witte, P.A.M.; Wolfender, J.-L.; Crawford, A.D. Zebrafish Bioassay-guided Microfractionation for the Rapid in vivo Identification of Pharmacologically Active Natural Products. Chim. Int. J. Chem. 2012, 66, 229–232. [Google Scholar] [CrossRef]
- Crawford, A.D.; Esguerra, C.V.; de Witte, P.A.M. Fishing for Drugs from Nature: Zebrafish as a Technology Platform for Natural Product Discovery. Planta Med 2008, 74, 624–632. [Google Scholar] [CrossRef] [Green Version]
- Zuberi, Z.; Eeza, M.N.H.; Matysik, J.; Berry, J.P.; Alia, A. NMR-Based Metabolic Profiles of Intact Zebrafish Embryos Exposed to Aflatoxin B1 Recapitulates Hepatotoxicity and Supports Possible Neurotoxicity. Toxins (Basel) 2019, 11, 258. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.C.; Hinsch, E.; Weber, G.; Freitas, S. Method of extraction and resolubilisation of pigments from Chlorociboria aeruginosa and Scytalidium cuboideum, two prolific spalting fungi. Coloration Technol. 2014, 130, 221–225. [Google Scholar] [CrossRef]
- Weber, G.; Boonloed, A.; Naas, K.M.; Koesdjojo, M.T.; Remcho, V.T.; Robinson, S.C. A method to stimulate production of extracellular pigments from wood-degrading fungi using a water carrier. Curr. Res. Environ. Appl. Mycol. 2016, 6, 218–230. [Google Scholar] [CrossRef]
- Vega Gutierrez, M.S.; Van Court, R.C.; Stone, D.W.; Konkler, M.J.; Groth, E.N.; Robinson, C.S. Relationship between Molarity and Color in the Crystal (Dramada’) produced by Scytalidium cuboideum, in Two Solvents. Molecules 2018, 23, 2581. [Google Scholar] [CrossRef] [Green Version]
- Giesbers, G.; Krueger, T.; Schenck, J.V.; Court, R.V.; Moore, J.; Fang, C.; Robinson, S.; Ostroverkhova, O. Fungi-derived xylindein: Effect of purity on optical and electronic properties. Mrs Adv. 2019, 4, 1769–1777. [Google Scholar] [CrossRef]
- Boonloed, A.; Weber, G.L.; Ramzy, K.M.; Dias, V.R.; Remcho, V.T. Centrifugal partition chromatography: A preparative tool for isolation and purification of xylindein from Chlorociboria aeruginosa. J. Chromatogr. A 2016, 1478, 19–25. [Google Scholar] [CrossRef]
- Vega Gutierrez, S.; Vega Gutierrez, P.; Godinez, A.; Pittis, L.; Huber, M.; Stanton, S.; Robinson, S. Feasibility of Coloring Bamboo with the Application of Natural and Extracted Fungal Pigments. Coatings 2016, 6, 37. [Google Scholar] [CrossRef] [Green Version]
- Hinsch, E.M.; Weber, G.; Chen, H.-L.; Robinson, S.C. Colorfastness of Extracted Wood-staining Fungal Pigments on Fabrics: A new potential for textile dyes. J. Text. Appar. Technol. Manag. 2015, 9. [Google Scholar] [CrossRef]
- Truong, L.; Harper, S.L.; Tanguay, R.L. Evaluation of embryotoxicity using the zebrafish model. Methods Mol. Biol. 2011, 691, 271–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandrell, D.; Truong, L.; Jephson, C.; Sarker, M.R.; Moore, A.; Lang, C.; Simonich, M.T.; Tanguay, R.L. Automated Zebrafish Chorion Removal and Single Embryo Placement: Optimizing Throughput of Zebrafish Developmental Toxicity Screens. J. Lab. Autom. 2012, 17, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Reif, D.M.; Truong, L.; Mandrell, D.; Marvel, S.; Zhang, G.; Tanguay, R.L. High-throughput characterization of chemical-associated embryonic behavioral changes predicts teratogenic outcomes. Arch. Toxicol. 2016, 90, 1459–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truong, L.; Zaikova, T.; Baldock, B.L.; Balik-Meisner, M.; To, K.; Reif, D.M.; Kennedy, Z.C.; Hutchison, J.E.; Tanguay, R.L. Systematic determination of the relationship between nanoparticle core diameter and toxicity for a series of structurally analogous gold nanoparticles in zebrafish. Nanotoxicology 2019, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Forest Products Lab. Extractives in Eastern Hardwoods—A Review; US Department of Agriculture: Masidon, WI, USA, 1979.
- He, Z.; Sleighter, R.L.; Hatcher, P.G.; Liu, S.; Wu, F.; Zou, H.; Olanya, O.M. Molecular level comparison of water extractives of maple and oak with negative and positive ion ESI FT-ICR mass spectrometry. J. Mass Spectrom. 2019, 54, 655–666. [Google Scholar] [CrossRef]
- VanderMolen, K.M.; Raja, H.A.; El-Elimat, T.; Oberlies, N.H. Evaluation of culture media for the production of secondary metabolites in a natural products screening program. Amb Express 2013, 3, 71. [Google Scholar] [CrossRef] [Green Version]
- Son, S.Y.; Lee, S.; Singh, D.; Lee, N.-R.; Lee, D.-Y.; Lee, C.H. Comprehensive Secondary Metabolite Profiling Toward Delineating the Solid and Submerged-State Fermentation of Aspergillus oryzae KCCM 12698. Front. Microbiol. 2018, 9, 1076. [Google Scholar] [CrossRef]
- Bode, H.; Bethe, B.; Hofs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. Chembiochem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Van Court, R.C.; Giesbers, G.; Ostroverkhova, O.; Robinson, C.S. Optimizing Xylindein from Chlorociboria spp. for (Opto)electronic Applications. Processes 2020, 8. [Google Scholar] [CrossRef]
- Tudor, D.; Margaritescu, S.; Sánchez-Ramírez, S.; Robinson, S.C.; Cooper, P.A.; Moncalvo, J.M. Morphological and molecular characterization of the two known North American Chlorociboria species and their anamorphs. Fungal Biol. 2014, 118, 732–742. [Google Scholar] [CrossRef]
- Petr, B.; Vojtech, A.; Ladislav, H.; Rene, K. Noteworthy Secondary Metabolites Naphthoquinones—Their Occurrence, Pharmacological Properties and Analysis. Curr. Pharm. Anal. 2009, 5, 47–68. [Google Scholar] [CrossRef]
- Kittakoop, P.; Punya, J.; Kongsaeree, P.; Lertwerawat, Y.; Jintasirikul, A.; Tanticharoen, M.; Thebtaranonth, Y. Bioactive naphthoquinones from Cordyceps unilateralis. Phytochemistry 1999, 52, 453–457. [Google Scholar] [CrossRef]
- Abe, F.R.; de Oliveira, D.P. Evaluation of apoptotic and necrotic cells of the natural dye erythrostominone. Toxicol. Lett. 2014, 229, S114. [Google Scholar] [CrossRef]
- Abe, F.R.; Soares, A.M.V.M.; Oliveira, D.P.d.; Gravato, C. Toxicity of dyes to zebrafish at the biochemical level: Cellular energy allocation and neurotoxicity. Environ. Pollut. 2018, 235, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Abe, F.R.; Mendonça, J.N.; Moraes, L.A.B.; Oliveira, G.A.R.d.; Gravato, C.; Soares, A.M.V.M.; Oliveira, D.P.d. Toxicological and behavioral responses as a tool to assess the effects of natural and synthetic dyes on zebrafish early life. Chemosphere 2017, 178, 282–290. [Google Scholar] [CrossRef] [PubMed]
- EPA (Ed.) Aniline Fact Sheet: Support Document (CAS No. 62-53-3); United States Evnironmental Protection Agency: Washington, DC, USA, 1994. [Google Scholar]
- Horie, Y.; Yamagishi, T.; Koshio, M.; Iguchi, T.; Tatarazako, N. Lethal and sublethal effects of aniline and chlorinated anilines on zebrafish embryos and larvae. J. Appl. Toxicol. 2017, 37, 836–841. [Google Scholar] [CrossRef]
- Zok, S.; Görge, G.; Kalsch, W.; Nagel, R. Bioconcentration, metabolism and toxicity of substituted anilines in the zebrafish (Brachydanio rerio). Sci. Total Environ. 1991, 109–110, 411–421. [Google Scholar] [CrossRef]
- Scientific, F. Material Safety Data Sheet Sodium chloride. Available online: https://fscimage.fishersci.com/msds/21105.htm (accessed on 17 February 2021).
- Verma, Y. Acute toxicity assessment of textile dyes and textile and dye industrial effluents using Daphnia magna bioassay. Toxicol. Ind. Health 2008, 24, 491–500. [Google Scholar] [CrossRef]
- Lopes, F.C.; Tichota, D.M.; Pereira, J.Q.; Segalin, J.; de Oliveira Rios, A.; Brandelli, A. Pigment Production by Filamentous Fungi on Agro-Industrial Byproducts: An Eco-Friendly Alternative. Appl. Biochem. Biotechnol. 2013, 171, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Palomino Agurto, E.M.; Vega Gutierrez, M.S.; Chen, H.-L.; Robinson, C.S. Wood-Rotting Fungal Pigments as Colorant Coatings on Oil-Based Textile Dyes. Coatings 2017, 7, 152. [Google Scholar] [CrossRef] [Green Version]
- Palomino Agurto, M.; Vega Gutierrez, S.; Van Court, R.; Chen, H.; Robinson, S. Oil-Based Fungal Pigment from Scytalidium cuboideum as a Textile Dye. J. Fungi 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Brun, N.R.; Wehrli, B.; Fent, K. Ecotoxicological assessment of solar cell leachates: Copper indium gallium selenide (CIGS) cells show higher activity than organic photovoltaic (OPV) cells. Sci. Total Environ. 2016, 543, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Cyrs, W.D.; Avens, H.J.; Capshaw, Z.A.; Kingsbury, R.A.; Sahmel, J.; Tvermoes, B.E. Landfill waste and recycling: Use of a screening-level risk assessment tool for end-of-life cadmium telluride (CdTe) thin-film photovoltaic (PV) panels. Energy Policy 2014, 68, 524–533. [Google Scholar] [CrossRef]
- Tsoutsos, T.; Frantzeskaki, N.; Gekas, V. Environmental impacts from the solar energy technologies. Energy Policy 2005, 33, 289–296. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almurshidi, B.H.; Van Court, R.C.; Vega Gutierrez, S.M.; Harper, S.; Harper, B.; Robinson, S.C. Preliminary Examination of the Toxicity of Spalting Fungal Pigments: A Comparison between Extraction Methods. J. Fungi 2021, 7, 155. https://doi.org/10.3390/jof7020155
Almurshidi BH, Van Court RC, Vega Gutierrez SM, Harper S, Harper B, Robinson SC. Preliminary Examination of the Toxicity of Spalting Fungal Pigments: A Comparison between Extraction Methods. Journal of Fungi. 2021; 7(2):155. https://doi.org/10.3390/jof7020155
Chicago/Turabian StyleAlmurshidi, Badria H., R.C. Van Court, Sarath M. Vega Gutierrez, Stacey Harper, Bryan Harper, and Seri C. Robinson. 2021. "Preliminary Examination of the Toxicity of Spalting Fungal Pigments: A Comparison between Extraction Methods" Journal of Fungi 7, no. 2: 155. https://doi.org/10.3390/jof7020155
APA StyleAlmurshidi, B. H., Van Court, R. C., Vega Gutierrez, S. M., Harper, S., Harper, B., & Robinson, S. C. (2021). Preliminary Examination of the Toxicity of Spalting Fungal Pigments: A Comparison between Extraction Methods. Journal of Fungi, 7(2), 155. https://doi.org/10.3390/jof7020155