

Unlocking the Potential of High-Amylose Starch for Gut Health: Not All Function the Same


Abstract
:1. Introduction
2. Structure and Unique Functional Properties of High-Amylose Starch
2.1. Multilevel Structure
2.2. Unique Functional Properties
3. Enzymatic Resistance of High-Amylose Starch

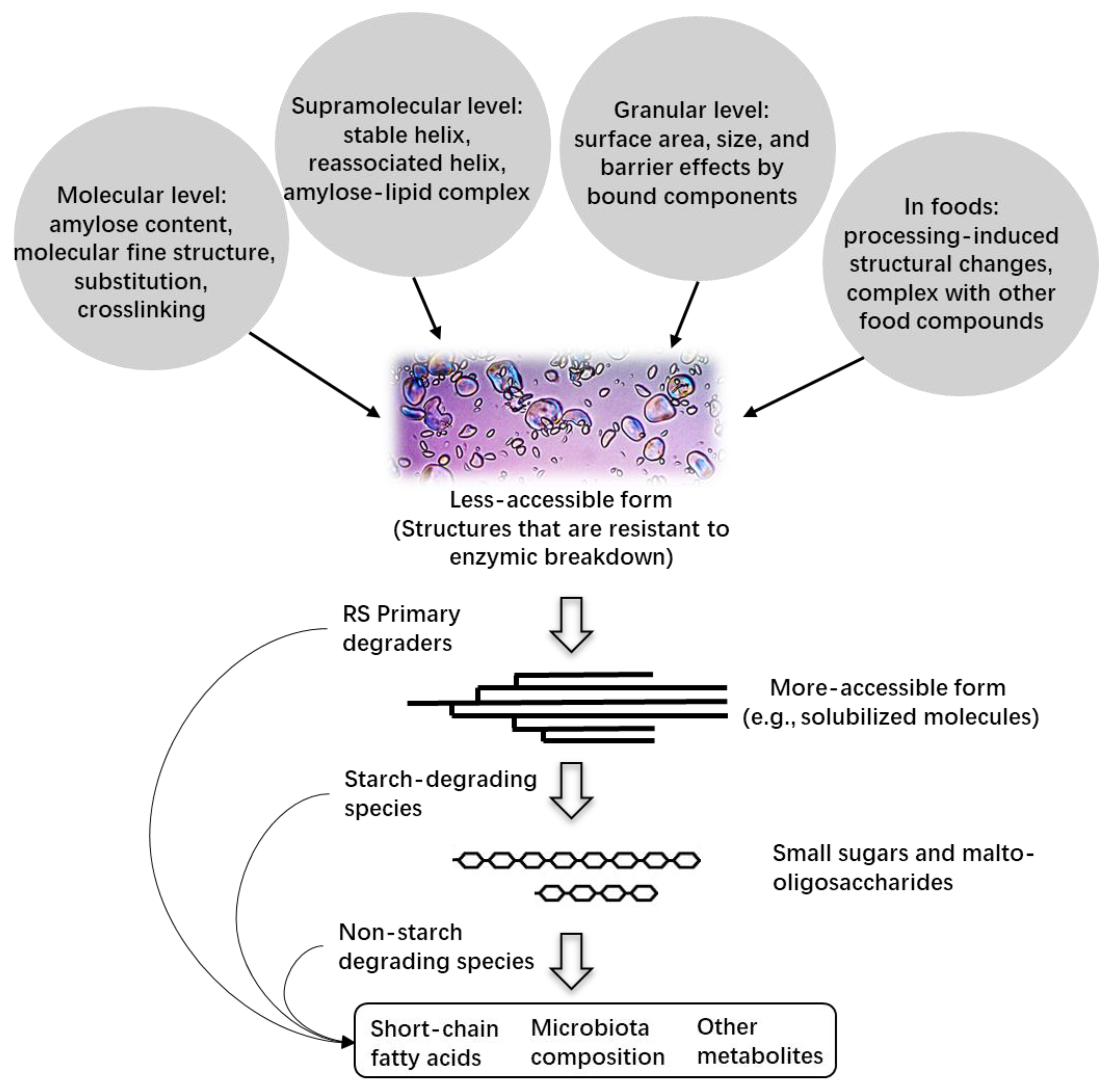{kind=link}
| Approach to Improve Amylose Content | Botanical Sources (Amylose Content, %) | Reference | Approach to Improve Amylose Content | Digestion Model Used | Resistant Starch Content (%) |
|---|---|---|---|---|---|
| Biological approach | Wheat (downregulated SS iIa: 34.0%→43.5%/ downregulated SBE iIa: 33.3%→57.8%); food processing into pasta | [43] | Downregulated SSIIa or SBE lla | In vitro model (pepsin, amyloglucosidase) | ↑1.33% (downregulated SS iIa)/↑7.15% (downregulated SBE iIa) |
| Wheat (bread wheat: 22.9%→55.7%/durum wheat: 24.4%→47.4%); milled | [26] | Downregulated SBE iIa | In vitro model (pancreatic alpha-amylase, amyloglucosidase) | ↑10.38% (bread wheat)/↑4.63% (durum wheat) | |
| Wheat (27.2%→84.2%) | [44] | Downregulated SBEs (IIa, IIb) | NA | ↑36.00% | |
| Wheat (27.9%→44.9%) | [45] | Downregulated SBEs (IIa, IIb) | Resistant starch kit | ↑5.50% | |
| Wheat (30.7%→50.0%) | [46] | Downregulated SBEs (IIa, IIb) | Resistant starch kit | ↑2.43% | |
| Wheat (23.0%→31.4%); cooked | [15] | Downregulated SBE (IIa) | In vitro model (pancreatic alpha-amylase, glucoamylase) | ↑ | |
| Wheat (32.3%→61.8%) | [47] | Targeted mutagenesis of SBEIIa | In vitro model (pancreatic alpha-amylase, amyloglucosidase) | ↑13.30% | |
| Rice (19.6%→41.2%); cooked | [20] | Downregulated SBE IIb | In vitro model (simulated oral phase, gastric phase, and intestinal phase) | ↑4.60% | |
| Rice (27.2%→64.8%) | [48] | Downregulated SBE I, SBE IIb | In vitro model (pancreatic alpha-amylase, amyloglucosidase) | ↑15.10% | |
| Chemical approach | Rice (30.6%) | [49] | Acid and heat-moisture treatments (citric acid/lactic acid/acetic acid) | In vitro model (alpha-amylase, amyloglucosidase) | ↑32.70% (citric acid) /↑28.80% (lactic acid) /↑26.70% (acetic acid) |
| Waxy rice (0.0%→30.3%); cooked (before pullulanase debranching) | [50] | Enzymatic modification (pullulanase) | In vitro model (pancreatic alpha-amylase, amyloglucosidase) | ↑28.28% | |
| Rice (enzymatic modification: 42.7%→49.2%/heat-moisture treatment: 42.7%→43.2%/acid treatment: 42.7%→43.9%) | [51] | Enzymatic modification (pullulanase)/heat-moisture treatment/acid treatment (citric acid) | In vitro model (pepsin, alpha-amylase, amyloglucosidase) | ↑6.26% (enzymatic modification)/↑2.52% (heat-moisture treatment)/↑10.77% (acid treatment) | |
| Maize (58.6%→58.5%) | [52] | Enzymatic modification (maltogenic alpha-amylase) | In vitro model (pancreatin, amyloglucosidase) | ↑1.00% | |
| Wheat (26.2%→35.6%) | [53] | Annealing (50 °C, 96 h, 1:3 w/v) | In vitro model (saccharifying enzyme, pancreatic alpha-amylase) | ↑4.61% | |
| Physical approach | Maize (85.0%→83.7%) | [54] | Annealing (45 °C, 72 h, 1:2 w/v) | In vitro model (pancreatic alpha-amylase, amyloglucosidase) | ↑0.60% |
| Maize (51.3%→49.1%) | [55] | Microwave (2450 MHz, 1.2 kW, 1 min) | In vitro model (pancreatin, amyloglucosidase) | ↑14.10% | |
| Barley (23.1%→23.4%) | [56] | Heat-moisture treatment (110 °C, 2 h, 30% moisture content) | In vitro model (pancreatin, amyloglucosidase) | ↑11.35% | |
| Quinoa (9.1%→16.4%) | [57] | Electron beam irradiation (8 kGy, 2 kGy/h) | In vitro model (pancreatin, amyloglucosidase) | ↑23.50% | |
| Potato (25.0%→28.8%) | [58] | Sonication (28 kHz, 300 W, 30 min, 25 °C) | In vitro model (pancreatic alpha-amylase, amyloglucosidase) | ↑ | |
| Lotus stem (41.9%→49.1%) | [59] | Sonication (20 kHz, 400 W, 35 min, 0 °C) | In vitro model (pancreatic alpha-amylase, amyloglucosidase) | ↑7.99% |
4. Colonic Fermentation of High-Amylose Starch and Nutritional Properties
4.1. In Vitro Studies and Animal Studies
| Fermentation Studies | Botanical Sources (Amylose Content, %) | Reference | Approach to Improve Amylose Content | Fermentation Models Used | Fermentation Properties (SCFA, Gas Production, and Microbiota Composition Changes) |
|---|---|---|---|---|---|
| Animal models | Maize (70%) | [84] | NA | In vivo fermentation model (7-week-old male rats) | Butyrate: ↑9.85 µmol per cecal content Acetate: ↑109.0 µmol per cecal content Propionate: ↑15.3 µmol per cecal content Total: ↑135.0 µmol per cecal content (cecal content, than low-amylose group) |
| Maize (63.3%) | [86] | NA | In vivo fermentation model (4-week-old male rats) | Butyrate: ↑10.1 µmol/g wet matter Acetate: ↑40.0 µmol/g wet matter Propionate: ↑10.3 µmol/g wet matter Total: ↑58.9 µmol/g wet matter Bacteroidetes ↑ (cecal content, than low-amylose group) | |
| Maize (85%) | [76] | NA | In vivo fermentation model (adult male rats) | Butyrate: ↑26.0 µmol Acetate: ↑225.0 µmol Propionate: ↑21.0 µmol Total: ↑276.0 µmol (cecal content, than low-amylose group) | |
| Maize (85%) | [78] | NA | In vivo fermentation model (4-week-old pigs) | Butyrate: ↑ Acetate: ↑ Propionate: ↑ Total: ↑ Total anaerobes↑, Lactobacillus ↑ (colonic digesta, than low-amylose group) | |
| Maize (80%) | [85] | NA | In vivo fermentation model (ileal-cannulated pigs) | Butyrate: ↑0.52 µmol/g wet matter Acetate: ↓0.7 µmol/g wet matter Propionate: ↑1.49 µmol/g wet matter Total: ↑8.5 µmol/g wet matter Bifidobacterium ↑ (feces, than low-amylose group) | |
| High-amylose maize | [82] | NA | In vivo fermentation model (male rats) | Butyrate: ↑ Acetate: ↑ Propionate: ↑ Total: ↑ Proteobacteria↓, Bacteroidetes↑ (feces, than low-amylose group) | |
| High-amylose maize | [93] | NA | In vivo fermentation model (adult male rats) | Butyrate: ↑ Acetate: ↑ Propionate: ↑ Total: ↑ (feces, than low-amylose group) | |
| High-amylose maize | [81] | NA | In vivo fermentation model (5-week-old male rats) | Butyrate: ↑3.0 μmol/g feces Acetate: ↑17.7 μmol/g feces Propionate: ↑7.0 μmol/g feces Total: ↑28.9 μmol/g feces (than low-amylose group) | |
| High-amylose maize | [83] | NA | In vivo fermentation model (adult male rats) | Butyrate: ↑24.9 µmol Acetate: ↑120.2 µmol Propionate: ↑38.9 µmol Total: ↑184.8 µmol (cecal content, than low-amylose group) | |
| High-amylose wheat/high-amylose maize | [79] | NA | In vivo fermentation model (male rats) | Butyrate: ↑9.0 µmol (wheat)/↑4.0 µmol (maize) Acetate: ↑21.0 µmol (wheat)/↑8.0 µmol (maize) Propionate: ↑12.0 µmol (wheat)/↑3.0 µmol (maize) Total: ↑44.0 µmol (wheat)/↑17.0 µmol (maize) (colonic digesta, than low-amylose group) | |
| Wheat (25.5%→74.4%) | [27] | Downregulated SBEs (IIa, IIb) | In vivo fermentation model (4-week-old male rats) | Butyrate: ↓2.8 mmol/kg Acetate: ↑5.0 mmol/kg Propionate: ↑3.9 mmol/kg Total: ↑4.5 mmol/kg (cecal digesta, than low-amylose group) | |
| Wheat (27.9%→44.9%) | [45] | Downregulated SBEs (IIa, IIb) | In vivo fermentation model (8-week-old rats) | Butyrate: ↑11.7 µmol Acetate: ↑79.8 µmol Propionate: ↑28.2 µmol Total: ↑119.7 µmol (cecal content, than wild-type starch) | |
| Rice (27.2%→64.8%) | [48] | Downregulated SBEs (I, IIb) | In vivo fermentation model (feeding rats for four weeks) | Butyrate: ↑11.9 μmol/g dry feces Acetate: ↑60.5 μmol/g dry feces Propionate: ↑68.1 μmol/g dry feces Total: ↑140.6 μmol/g dry feces (feces, than wild-type starch) | |
| Barley (41%) | [80] | NA | In vivo fermentation model (4-week-old pigs) | Butyrate: ↓4% Acetate: ↓6.4% Propionate: ↑7.9% Total: ↑19 mMol/kg digesta sample NH3: ↑1 mMol/kg digesta sample (colonic digesta, than low-amylose group) | |
| High-amylose barley | [77] | NA | In vivo fermentation model (adult male rats) | Butyrate: ↑12.0 µmol Acetate: ↑142.0 µmol Propionate: ↑9.0 µmol Total: ↑161.0 µmol (feces, than low-amylose group) | |
| In vitro models | High-amylose maize | [72] | Propionylated | In vitro fermentation model (bacteroides-dominated enterotype inocula) | Butyrate: ↓1.0 mM/50 mg dry feces Acetate: ↑0.2 mM/50 mg dry feces Propionate: ↑0.7 mM/50 mg dry feces Total: ↑2.4 mM/50 mg dry feces Bacteroidetes ↑ Gas production → (than unmodified starch) |
| High-amylose maize; Cooked | [71] | NA | In vitro fermentation model (inoculum from human stool samples) | Butyrate: ↑ Acetate: ↑ Propionate: ↑ Total: ↑ (than low-amylose group) | |
| High-amylose maize | [70] | Debranched (pullulanase), recrystallization combined with HMT | In vitro fermentation model (inoculum from human stool samples) | Butyrate: ↑2.4 mM Acetate: ↑1.8 mM Propionate: ↑2.7 mM Total: ↑7.4 mM (fermentation for 24 h, than unmodified starch) | |
| High-amylose maize; cooked, recrystallized, and amylase digested | [75] | NA | In vitro fermentation model (inoculum from pig fecal samples) | Butyrate: → Acetate: → Propionate: → Total: ↓0.6 mmol/g dry matter DMCV: ↑14 mL, Rmax: ↓0.8 mL/h NH3 production: ↓ (fermentation for 146 h, than cooked and recrystallized group) | |
| Maize (69.8%) | [73] | Propionylated | In vitro fermentation model (inoculum from human stool samples) | Butyrate: ↓ Acetate: ↑ Propionate: ↑ Total: ↑ Firmicutes ↑, Bacteroidetes: ↓ Gas production → (fermentation for 24 h, than unmodified starch) | |
| Maize (65.0%) | [74] | Chemical cross-linking | In vitro fermentation model (inoculum from human stool samples) | Butyrate: ↓ Acetate: ↑ Propionate: → Total: ↓ Firmicutes/Bacteroidetes ratio: ↓ Gas production: ↓ (fermentation for 24 h, than unmodified starch) | |
| Wheat (32%→84%) | [67] | Downregulated SBEs (IIa, IIb) | In vitro fermentation model (using wheat-based foods and inoculum from human stool samples) | Butyrate: ↓ Acetate: ↓ Propionate: ↑ Total: ↓ DMCV: ↓10 mL/gDM, Rmax: ↓ (fermentation for 48 h, than wild-type starch) | |
| Wheat (37%→93%) | [61] | Downregulated SBEs (IIa, IIb) | In vitro fermentation model (inoculum from human stool samples) | Butyrate: ↓ Acetate: ↓ Propionate: ↓ Total: ↓ DMCV: ↓55 mL/g DM, Rmax: ↓6.3 mL/h, NH4+ production: no difference (fermentation for 72 h, than wild-type starch) |
4.2. Dietary Intervention Studies
| Botanical Sources | Reference | Modifications or Food Processing | Study Design | Results | |
|---|---|---|---|---|---|
| Wheat | High-amylose wheat | [94] | Bread/biscuits with low-amylose wheat or low-amylose wheat refined, high-amylose wheat or high-amylose wheat refined | Healthy subjects (n = 80), random, double-blinded, 4-arm parallel | Fecal butyrate excretion: ↑38% (p < 0.05, refined high-amylose wheat vs. refined low-amylose wheat) Abundance of fecal SCFA-producing bacteria: ↑Roseburia inulinivorans (p < 0.001) of refined high-amylose wheat than at baseline (week 0) |
| High-amylose maize flour (amylose content = 67%) | [105] | Cookies with high-amylose maize flour, low-amylose maize flour, or 100% wheat flour | Healthy men (n = 30), random, double-blinded | Cumulative postprandial glycemic AUC (0 to 200 min): ↓0.18 mmol/L (p < 0.05) Cumulative blood glucose AUC (0 to 200 min): ↓75.53 mmol×min/L (p < 0.05) (than 100% wheat flour) | |
| High-amylose wheat (amylose content = 74.3%) | [100] | Bread containing low-amylose wheat or low-amylose wheat refined, high-amylose wheat or high-amylose wheat refined | Healthy subjects (n = 20), random, double-blinded | Postprandial glycemic AUC: ↓25 mmol/L×3 h (p < 0.001) Plasma glucose concentration: ↓33% Glycemic AUC response: ↓39% (p < 0.0001) Insulinemic AUC response: ↓24–30% (p < 0.05) (than low-amylose group) Whole-meal or refined flour did not affect the glycemic, insulinemic, or incretin response | |
| Maize | High-amylose maize (amylose content = 33%) | [96] | Muffins with low or high (RS2 20 g) amylose maize starch | Hypertriglyceridemia subjects (n = 23), random | Postprandial glucose area: ↓0.25 mmol × min/L SCFA concentrations in fecal water: ↑32% (p < 0.001) Overall postprandial plasma insulin concentration: ↓17% Postprandial insulin area: ↓62 pmol × min/L (p < 0.01) Frequency of bowel actions: ↑0.2 actions/day (than low-amylose group) |
| High-amylose maize starch | [95] | Supplements containing HAMS (RS 30 g/day) or low-fiber content | Healthy subjects (n = 24), random | Fecal bulk: ↑22 g/day (p < 0.001) Fecal SCFA output: ↑2.32 mmol/day Fecal SCFA concentration: ↑5.3 mmol/L Fecal butyrate: SCFA ratio significantly 31% (p = 0.035) (than low-fiber supplement) | |
| High-amylose maize starch | [102] | Breakfast with amylopectin starch plus cellulose, amylopectin starch plus lactulose, or high-amylose starch plus cellulose | Healthy subjects (n = 10), random, single-blinded | PPGR: ↓ Glucose tolerance: ↑ Non-esterified fatty acids concentrations: ↑ (high-amylose starch plus cellulose vs. amylopectin starch plus cellulose group) | |
| High-amylose maize starch | [116] | Sachets containing waxy maize starch (Amioca) or HAMS (RS2 30 g/day) | Healthy subjects (n = 10), random, single-blinded | SCFA concentrations (acetate and propionate): ↑ (not colonic SCFAs) Plasma insulin response: ↓ No effect on Postprandial glycemic AUC, HOMA, and FBG than waxy maize starch | |
| High-RS maize starch | [111] | Bread with or without HAMS | Healthy subjects (n = 15), random | Blood glucose incremental AUC: ↓44.4 mmol × min/L Insulin incremental AUC: ↓4.8 pmol × min/L (p < 0.05) Colonic fermentation in the late postprandial phase: ↑23.3 ± 4.5 ppm (breath H2) No effect on PPGR (than white wheat flour bread) | |
| High-amylose maize starch (resistant starch content = 60%) | [112] | A test breakfast and lunch containing either 48 g RS or not | Healthy males (n = 10), random, single-blinded, cross-over | PPGR: ↓ AUC C-peptide: ↑26,457 nmol/L 300 min AUC Insulin: ↑40,714 pmol/L 300 min (than breakfast and lunch without RS) | |
| High-amylose maize starch | [103] | Chinese steamed bun formulation containing either HAMS or not | Healthy females (n = 15), random, single-blinded | PPGR: ↓80.9 mmol × min/L (p = 0.004) Mean GI: ↓30.07 Incremental postprandial glycemic AUC values: ↓ (than wheat flour without HAMS) | |
| High-amylose maize | [110] | Sachets containing waxy maize starch (Amioca) or HAMS (RS2 15 or 30 g/day) | Obese subjects (n = 33), random, double-blinded | FSIVGTT: ↑in men (n = 11) Fecal SCFA output: ↑ No effect on HOMA and FBG (than waxy maize starch) | |
| High-amylose maize starch | [97] | Bread with or without HAMS (RS2 12 g/day) | Obese subjects (n = 15), random, subject-blinded | Fecal acetate: ↓6.87 μmol/L Fecal propionate: ↑0.82 μmol/L Fecal butyrate: ↑0.08 μmol/L (than bread without HAMS) | |
| High-amylose maize | [108] | Sachets containing waxy maize starch or HAMS (RS2 40 g/day) | Insulin-resistant subjects (n = 15), random, controlled | HOMA for insulin resistance: ↓ (p = 0.029) Fasting insulin: ↓21 pmol/liter (p = 0.041) FBG: ↓(p = 0.017) Postprandial glucose disposal: ↑65% (than waxy maize starch) | |
| High-amylose maize | [113] | Cookies containing waxy maize starch or HAMS (RS2 15 or 30 g/day) | Healthy and non-diabetic women (n = 51; 23 women completed all 3 arms), random, placebo-controlled, double-blinded, cross-over | Insulin resistant group (n = 14): insulin sensitivity after consuming HAMS 30 g/day ↑(than HAMS 15 g/day); Insulin sensitive group (n = 9): no significant difference (p > 0.05) Both groups: no significant differences in fasting glucose or insulin after consuming HAMS 15 or 30 g/day (than waxy maize starch) | |
| High-amylose maize starch | [114] | Bagel containing only hard wheat flour or 60% substitution of HAMS (RS2 25 g/day) | Subjects at high risk of type 2 diabetes (n = 24), random, double-blinded | HOMA for insulin resistance: ↓ Insulin incremental AUC: ↓18.9% (p = 0.04) (than hard wheat flour) | |
| High-amylose maize | [106] | Muffins with or without HAM (RS2 30 or 0 g/day) | Overweight, healthy adults (n = 18), randomized-controlled, parallel-arm, double-blinded | Postprandial glycemic AUC: ↓ Glucose homeostasis: ↑ Insulin change not observed (than muffins without HAM) | |
| High-amylose maize starch | [107] | Muffin top containing modified HAMS (RS4) or not | Healthy adults (n = 12), random, double-blinded, controlled | Postprandial glycemic AUC: ↓33% (p = 0.037) Maximum glucose concentration: ↓8% Postprandial serum insulin incremental AUC: ↓38% (p < 0.001) (than control) | |
| High-amylose maize | [101] | Sachets containing waxy maize starch or HAMS (RS2 40 g/ day) | Insulin-resistant, obese subjects (n = 12), random, single-blinded | Insulin and C-peptide concentrations: ↑ Glucose effectiveness:↓0.01/min (p = 0.06) (than waxy maize starch) | |
| High-amylose maize starch | [99] | Soups with 50 g maltodextrin, whole grain (RS 27 g/day), high-amylose (RS 23 g/day), regular cornstarch, or no added starch | Healthy men (n = 17), random | Postprandial glycemic AUC: ↓ (than soups without added starch) | |
| Barley | High-amylose maize | [104] | Breakfast and lunch containing barley flour (more HAM) or white wheat flour (without HAM) | Healthy women (n = 14), random, single-blinded | Postprandial glycemic AUC: ↓22% AUC of insulin: ↓32% No effect on HOMA and FBG (than white wheat flour) |
| High-starch amylose (amylose content = 42%) | [109] | Barley tortillas varying in fiber and/or starch composition | Healthy adults (n = 12), random, double-blinded | No significant difference in GI between the low-amylose and high-amylose groups No difference in the glucose iAUC or percentage change at 30 min between the low-amylose and high-amylose groups Insulin concentrations at 15–60 min: ↑ all groups | |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.N.; Akerman, A.P.; Mann, J. Dietary fibre and whole grains in diabetes management: Systematic review and meta-analyses. PLoS Med. 2020, 17, e1003053. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Te Morenga, L. Carbohydrate quality and human health: A series of systematic reviews and meta-analyses. Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Ding, M.; Sampson, L.; Willett, W.C.; Manson, J.E.; Wang, M.; Rosner, B.; Hu, F.B.; Sun, Q. Intake of whole grain foods and risk of type 2 diabetes: Results from three prospective cohort studies. BMJ 2020, 370, m2206. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-T.; Li, Z.; Fox, G.P.; Gidley, M.J.; Dhital, S. Protein-starch matrix plays a key role in enzymic digestion of high-amylose wheat noodle. Food Chem. 2021, 336, 127719. [Google Scholar] [CrossRef]
- Reyniers, S.; De Brier, N.; Ooms, N.; Matthijs, S.; Piovesan, A.; Verboven, P.; Brijs, K.; Gilbert, R.G.; Delcour, J.A. Amylose molecular fine structure dictates water–oil dynamics during deep-frying and the caloric density of potato crisps. Nat. Food 2020, 1, 736–745. [Google Scholar] [CrossRef]
- Li, C.; Dhital, S.; Gidley, M.J. High-amylose wheat bread with reduced in vitro digestion rate and enhanced resistant starch content. Food Hydrocoll. 2021, 123, 107181. [Google Scholar] [CrossRef]
- Seung, D. Amylose in starch: Towards an understanding of biosynthesis, structure and function. New Phytol. 2020, 228, 1490–1504. [Google Scholar] [CrossRef]
- Li, H.-T.; Gidley, M.J.; Dhital, S. High-amylose starches to bridge the “fiber gap”: Development, structure, and nutritional functionality. Compr. Rev. Food Sci. Food Saf. 2019, 18, 362–379. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Tan, H.; Zhang, C.; Li, Q.; Liu, Q. Starch biosynthesis in cereal endosperms: An updated review over the last decade. Plant Commun. 2021, 2, 100237. [Google Scholar] [CrossRef]
- Birt, D.F.; Boylston, T.; Hendrich, S.; Jane, J.-L.; Hollis, J.; Li, L.; McClelland, J.; Moore, S.; Phillips, G.J.; Rowling, M.; et al. Resistant starch: Promise for improving human health. Adv. Nutr. 2013, 4, 587–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bird, A.R.; Regina, A. High amylose wheat: A platform for delivering human health benefits. J. Cereal Sci. 2018, 82, 99–105. [Google Scholar] [CrossRef]
- Tizzotti, M.J.; Sweedman, M.C.; Tang, D.; Schaefer, C.; Gilbert, R.G. New 1H NMR Procedure for the Characterization of Native and Modified Food-Grade Starches. J. Agric. Food Chem. 2011, 59, 6913–6919. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.-C.; Capitani, T.; Trzasko, P.; Jeffcoat, R. Molecular structure of a low-amylopectin starch and other high-amylose maize starches. J. Cereal Sci. 1998, 27, 289–299. [Google Scholar] [CrossRef]
- Shimbata, T.; Ai, Y.; Fujita, M.; Inokuma, T.; Vrinten, P.; Sunohara, A.; Saito, M.; Takiya, T.; Jane, J.-L.; Nakamura, T. Effects of homoeologous wheat starch synthase IIa genes on starch properties. J. Agric. Food Chem. 2012, 60, 12004–12010. [Google Scholar] [CrossRef] [PubMed]
- Vilaplana, F.; Meng, D.; Hasjim, J.; Gilbert, R.G. Two-dimensional macromolecular distributions reveal detailed architectural features in high-amylose starches. Carbohydr. Polym. 2014, 113, 539–551. [Google Scholar] [CrossRef] [Green Version]
- Jane, J.-L. Current understanding on starch granule structures. J. Appl. Glycosci. 2006, 53, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Li, H.-T.; Dhital, S.; Flanagan, B.M.; Mata, J.; Gilbert, E.P.; Gidley, M.J. High-amylose wheat and maize starches have distinctly different granule organization and annealing behaviour: A key role for chain mobility. Food Hydrocoll. 2020, 105, 105820. [Google Scholar] [CrossRef]
- Wang, T.; Bogracheva, T.; Hedley, C. Starch: As simple as A, B, C? J. Exp. Bot. 1998, 49, 481–502. [Google Scholar] [CrossRef] [Green Version]
- Butardo, V.M.; Fitzgerald, M.A.; Bird, A.R.; Gidley, M.J.; Flanagan, B.M.; Larroque, O.; Resurreccion, A.P.; Laidlaw, H.K.C.; Jobling, S.A.; Morell, M.K.; et al. Impact of down-regulation of starch branching enzyme IIb in rice by artificial micro RNA- and hairpin RNA- mediated RNA silencing. J. Exp. Bot. 2011, 62, 4927–4941. [Google Scholar] [CrossRef]
- Huang, J.; Shang, Z.; Man, J.; Liu, Q.; Zhu, C.; Wei, C. Comparison of molecular structures and functional properties of high-amylose starches from rice transgenic line and commercial maize. Food Hydrocoll. 2015, 46, 172–179. [Google Scholar] [CrossRef]
- Regina, A.; Blazek, J.; Gilbert, E.; Flanagan, B.M.; Gidley, M.J.; Cavanagh, C.; Ral, J.-P.; Larroque, O.; Bird, A.R.; Li, Z.; et al. Differential effects of genetically distinct mechanisms of elevating amylose on barley starch characteristics. Carbohydr. Polym. 2012, 89, 979–991. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-T.; Dhital, S.; Slade, A.J.; Yu, W.; Gilbert, R.G.; Gidley, M.J. Altering starch branching enzymes in wheat generates high-amylose starch with novel molecular structure and functional properties. Food Hydrocoll. 2019, 92, 51–59. [Google Scholar] [CrossRef]
- Yamamori, M.; Fujita, S.; Hayakawa, K.; Matsuki, J.; Yasui, T. Genetic elimination of a starch granule protein, SGP-1, of wheat generates an altered starch with apparent high amylose. Int. J. Plant Breed. Res. 2000, 101, 21–29. [Google Scholar] [CrossRef]
- Blazek, J.; Salman, H.; Rubio, A.L.; Gilbert, E.; Hanley, T.; Copeland, L. Structural characterization of wheat starch granules differing in amylose content and functional characteristics. Carbohydr. Polym. 2009, 75, 705–711. [Google Scholar] [CrossRef] [Green Version]
- Slade, A.J.; McGuire, C.; Loeffler, D.; Mullenberg, J.; Skinner, W.; Fazio, G.; Holm, A.; Brandt, K.M.; Steine, M.N.; Goodstal, J.F.; et al. Development of high amylose wheat through TILLING. BMC Plant Biol. 2012, 12, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Regina, A.; Bird, A.; Topping, D.; Bowden, S.; Freeman, J.; Barsby, T.; Kosar-Hashemi, B.; Li, Z.; Rahman, S.; Morell, M. High-amylose wheat generated by RNA interference improves indices of large-bowel health in rats. Proc. Natl. Acad. Sci. USA 2006, 103, 3546–3551. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Horner, H.T.; Pepper, T.M.; Blanco, M.; Campbell, M.; Jane, J.-L. Formation of elongated starch granules in high-amylose maize. Carbohydr. Polym. 2010, 80, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.; Qin, F.; Zhu, L.; Zhou, W.; Chen, Y.; Wang, Y.; Gu, M.; Liu, Q. Microstructure and ultrastructure of high-amylose rice resistant starch granules modified by antisense RNA inhibition of starch branching enzyme. J. Agric. Food Chem. 2010, 58, 1224–1232. [Google Scholar] [CrossRef]
- Li, H.-T.; Sartika, R.S.; Kerr, E.D.; Schulz, B.L.; Gidley, M.J.; Dhital, S. Starch granular protein of high-amylose wheat gives innate resistance to amylolysis. Food Chem. 2020, 330, 127328. [Google Scholar] [CrossRef]
- Ye, X.; Zhang, Y.; Qiu, C.; Corke, H.; Sui, Z. Extraction and characterization of starch granule-associated proteins from rice that affect in vitro starch digestibility. Food Chem. 2019, 276, 754–760. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.K.; Blazek, J.; Flanagan, B.M.; Dhital, S.; Larroque, O.; Morell, M.K.; Gilbert, E.P.; Gidley, M.J. Molecular, mesoscopic and microscopic structure evolution during amylase digestion of extruded maize and high amylose maize starches. Carbohydr. Polym. 2015, 118, 224–234. [Google Scholar] [CrossRef]
- Dhital, S.; Shrestha, A.K.; Gidley, M.J. Relationship between granule size and in vitro digestibility of maize and potato starches. Carbohydr. Polym. 2010, 82, 480–488. [Google Scholar] [CrossRef]
- Gallant, D.J.; Bouchet, B.; Buléon, A.; Pérez, S. Physical characteristics of starch granules and susceptibility to enzymatic degradation. Eur. J. Clin. Nutr. 1992, 46, S3–S16. [Google Scholar]
- Zhang, G.; Ao, Z.; Hamaker, B. Slow digestion property of native cereal starches. Biomacromolecules 2006, 7, 3252–3258. [Google Scholar] [CrossRef] [PubMed]
- Jane, J.-L.; Chen, Y.Y.; Lee, L.F.; McPherson, A.E.; Wong, K.S.; Radosavljevic, M.; Kasemsuwan, T. Effects of amylopectin branch chain length and amylose content on the gelatinization and pasting properties of starch. Cereal Chem. 1999, 76, 629–637. [Google Scholar] [CrossRef]
- Zhang, G.; Ao, Z.; Hamaker, B.R. Nutritional property of endosperm starches from maize mutants: A parabolic relationship between slowly digestible starch and amylopectin fine structure. J. Agric. Food Chem. 2008, 56, 4686–4694. [Google Scholar] [CrossRef]
- Li, H.-T.; Dhital, S.; Flanagan, B.M.; Mata, J.; Gilbert, E.P.; Gilbert, R.G.; Gidley, M.J. Amorphous packing of amylose and elongated branches linked to the enzymatic resistance of high-amylose wheat starch granules. Carbohydr. Polym. 2022, 295, 119871. [Google Scholar] [CrossRef]
- Shrestha, A.K.; Blazek, J.; Flanagan, B.M.; Dhital, S.; Larroque, O.; Morell, M.K.; Gilbert, E.P.; Gidley, M.J. Molecular, mesoscopic and microscopic structure evolution during amylase digestion of maize starch granules. Carbohydr. Polym. 2012, 90, 23–33. [Google Scholar] [CrossRef]
- Tester, R.F.; Qi, X.; Karkalas, J. Hydrolysis of native starches with amylases. Anim. Feed Sci. Technol. 2006, 130, 39–54. [Google Scholar] [CrossRef]
- Gidley, M.J. Factors affecting the crystalline type (A-C) of native starches and model compounds: A rationalisation of observed effects in terms of polymorphic structures. Carbohydr. Res. 1987, 161, 301–304. [Google Scholar] [CrossRef]
- Wang, S.; Li, C.; Copeland, L.; Niu, Q.; Wang, S. Starch Retrogradation: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2015, 14, 568–585. [Google Scholar] [CrossRef]
- Sissons, M.; Sestili, F.; Botticella, E.; Masci, S.; Lafiandra, D. Can manipulation of Durum wheat amylose content reduce the glycaemic index of Spaghetti? Foods 2020, 9, 693. [Google Scholar] [CrossRef]
- Regina, A.; Berbezy, P.; Kosar-Hashemi, B.; Li, S.; Cmiel, M.; Larroque, O.; Bird, A.R.; Swain, S.M.; Cavanagh, C.; Jobling, S.A.; et al. A genetic strategy generating wheat with very high amylose content. Plant Biotechnol. J. 2015, 13, 1276–1286. [Google Scholar] [CrossRef]
- Hazard, B.; Zhang, X.; Naemeh, M.; Hamilton, M.K.; Rust, B.; Raybould, H.E.; Newman, J.W.; Martin, R.; Dubcovsky, J. Mutations in durum wheat genes affect grain yield components, quality, and fermentation responses in rats. Crop Sci. 2015, 55, 2813. [Google Scholar] [CrossRef] [Green Version]
- Schonhofen, A.; Hazard, B.; Zhang, X.; Dubcovsky, J. Registration of common wheat germplasm with mutations in SBEII genes conferring increased grain amylose and resistant starch content. J. Plant Regist. 2016, 10, 200–205. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Jiao, G.; Sun, Y.; Chen, J.; Zhong, Y.; Yan, L.; Jiang, D.; Ma, Y.; Xia, L. Modification of starch composition, structure and properties through editing of TaSBEIIa in both winter and spring wheat varieties by CRISPR/Cas9. Plant Biotechnol. 2020, 19, 937–951. [Google Scholar] [CrossRef]
- Zhu, L.; Gu, M.; Meng, X.; Cheung, S.C.K.; Yu, H.; Huang, J.; Sun, Y.; Shi, Y.; Liu, Q. High-amylose rice improves indices of animal health in normal and diabetic rats. Plant Biotechnol. J. 2012, 10, 353–362. [Google Scholar] [CrossRef]
- Hung, P.V.; Vien, N.L.; Lan Phi, N.T. Resistant starch improvement of rice starches under a combination of acid and heat-moisture treatments. Food Chem. 2016, 191, 67–73. [Google Scholar] [CrossRef]
- Shi, M.-M.; Gao, Q.-Y. Physicochemical properties, structure and in vitro digestion of resistant starch from waxy rice starch. Carbohydr. Polym. 2011, 84, 1151–1157. [Google Scholar] [CrossRef]
- Bao, J.; Zhou, X.; Hu, Y.; Zhang, Z. Resistant starch content and physicochemical properties of non-waxy rice starches modified by pullulanase, heat-moisture treatment, and citric acid. J. Cereal Sci. 2022, 105, 103472. [Google Scholar] [CrossRef]
- Li, J.; Kong, X.; Ai, Y. Modification of granular waxy, normal and high-amylose maize starches by maltogenic α-amylase to improve functionality. Carbohydr. Polym. 2022, 290, 119503. [Google Scholar] [CrossRef]
- Su, C.; Saleh, A.S.; Zhang, B.; Zhao, K.; Ge, X.; Zhang, Q.; Li, W. Changes in structural, physicochemical, and digestive properties of normal and waxy wheat starch during repeated and continuous annealing. Carbohydr. Polym. 2020, 247, 116675. [Google Scholar] [CrossRef]
- Wang, S.; Wang, J.; Yu, J.; Wang, S. A comparative study of annealing of waxy, normal and high-amylose maize starches: The role of amylose molecules. Food Chem. 2014, 164, 332–338. [Google Scholar] [CrossRef]
- Zhong, Y.; Liang, W.; Pu, H.; Blennow, A.; Liu, X.; Guo, D. Short-time microwave treatment affects the multi-scale structure and digestive properties of high-amylose maize starch. Int. J. Biol. Macromol. 2019, 137, 870–877. [Google Scholar] [CrossRef]
- Liu, K.; Zhang, B.; Chen, L.; Li, X.; Zheng, B. Hierarchical structure and physicochemical properties of highland barley starch following heat moisture treatment. Food Chem. 2019, 271, 102–108. [Google Scholar] [CrossRef]
- Du, Z.; Li, Y.; Luo, X.; Xing, J.; Zhang, Q.; Wang, R.; Wang, L.; Chen, Z. Effects of electron beam irradiation on the physicochemical properties of quinoa and starch microstructure. Starch Stärke 2020, 72, 1900178. [Google Scholar] [CrossRef]
- Xiao, Y.; Wu, X.; Zhang, B.; Luo, F.; Lin, Q.; Ding, Y. Understanding the aggregation structure, digestive and rheological properties of corn, potato, and pea starches modified by ultrasonic frequency. Int. J. Biol. Macromol. 2021, 189, 1008–1019. [Google Scholar] [CrossRef]
- Noor, N.; Gani, A.; Jhan, F.; Jenno, J.; Dar, M.A. Resistant starch type 2 from lotus stem: Ultrasonic effect on physical and nutraceutical properties. Ultrason. Sonochem. 2021, 76, 105655. [Google Scholar] [CrossRef]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef] [Green Version]
- Bui, A.T.; Williams, B.A.; Hoedt, E.C.; Morrison, M.; Mikkelsen, D.; Gidley, M.J. High amylose wheat starch structures display unique fermentability characteristics, microbial community shifts and enzyme degradation profiles. Food Funct. 2020, 11, 5635–5646. [Google Scholar] [CrossRef] [PubMed]
- DeMartino, P.; Cockburn, D.W. Resistant starch: Impact on the gut microbiome and health. Curr. Opin. Biotechnol. 2020, 61, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, F.M.; Photenhauer, A.L.; Pollet, R.M.; Brown, H.A.; Koropatkin, N.M. Starch digestion by gut bacteria: Crowdsourcing for carbs. Trends Microbiol. 2020, 28, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.K.; Rossi, M.; Bajka, B.; Whelan, K. Dietary fibre in gastrointestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 101–116. [Google Scholar] [CrossRef]
- Cantu-Jungles, T.M.; Hamaker, B.R. New view on dietary fiber selection for predictable shifts in gut microbiota. mBio 2020, 11, 2179–2188. [Google Scholar] [CrossRef] [Green Version]
- Hamaker, B.R.; Tuncil, Y.E. A perspective on the complexity of dietary fiber structures and their potential effect on the gut microbiota. J. Mol. Biol. 2014, 426, 3838–3850. [Google Scholar] [CrossRef]
- Bui, A.T.; Williams, B.A.; Murtaza, N.; Lisle, A.; Mikkelsen, D.; Morrison, M.; Gidley, M.J. Wheat-based food form has a greater effect than amylose content on fermentation outcomes and microbial community shifts in an in vitro fermentation model. Food Hydrocoll. 2020, 114, 106560. [Google Scholar] [CrossRef]
- Williams, B.A.; Verstegen, M.W.A.; Tamminga, S. Fermentation in the large intestine of single-stomached animals and its relationship to animal health. Nutr. Res. Rev. 2001, 14, 207–228. [Google Scholar] [CrossRef]
- Cummings, J.H.; Macfarlane, G.T. The control and consequences of bacterial fermentation in the human colon. J. Appl. Bacteriol. 1991, 70, 443–459. [Google Scholar] [CrossRef]
- Chang, R.; Jin, Z.; Lu, H.; Qiu, L.; Sun, C.; Tian, Y. Type III resistant starch prepared from debranched starch: Structural changes under simulated saliva, gastric, and intestinal conditions and the impact on short-chain fatty acid production. J. Agric. Food Chem. 2021, 69, 2595–2602. [Google Scholar] [CrossRef]
- Liu, Y.; Matheyambath, A.C.; Polic, I.I.; LaPointe, G. Differential fermentation of raw and processed high-amylose and waxy maize starches in the Simulator of the Human Intestinal Microbial Ecosystem (SHIME®). J. Funct. Foods 2021, 86, 104735. [Google Scholar] [CrossRef]
- Xie, Z.; Ding, L.; Huang, Q.; Fu, X.; Liu, F.; Dhital, S.; Zhang, B. In vitro colonic fermentation profiles and microbial responses of propionylated high-amylose maize starch by individual Bacteroides-dominated enterotype inocula. Food Res. Int. 2021, 144, 110317. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Wang, S.; Wang, Z.; Fu, X.; Huang, Q.; Yuan, Y.; Wang, K.; Zhang, B. In vitro fecal fermentation of propionylated high-amylose maize starch and its impact on gut microbiota. Carbohydr. Polym. 2019, 223, 115069. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, B.; Chen, T.; Li, C.; Fu, X.; Huang, Q. Chemical cross-linking controls in vitro fecal fermentation rate of high-amylose maize starches and regulates gut microbiota composition. J. Agric. Food Chem. 2019, 67, 13728–13736. [Google Scholar] [CrossRef]
- Warren, F.J.; Fukuma, N.M.; Mikkelsen, D.; Flanagan, B.M.; Williams, B.A.; Lisle, A.T.; Cuiv, P.O.; Morrison, M.; Gidley, M.J. Food starch structure impacts gut microbiome composition. Msphere 2018, 3, e00086-18. [Google Scholar] [CrossRef] [Green Version]
- Bird, A.R.; Brown, I.L.; Topping, D.L. Low and high amylose maize starches acetylated by a commercial or a laboratory process both deliver acetate to the large bowel of rats. Food Hydrocoll. 2006, 20, 1135–1140. [Google Scholar] [CrossRef]
- Bird, A.R.; Flory, C.; Davies, D.A.; Usher, S.; Topping, D.L. A novel barley cultivar (Himalaya 292) with a specific gene mutation in starch synthase lla raises large bowel starch and short-chain fatty acids in rats. J. Nutr. 2004, 134, 831–835. [Google Scholar] [CrossRef] [Green Version]
- Bird, A.R.; Vuaran, M.; Brown, I.; Topping, D.L. Two high-amylose maize starches with different amounts of resistant starch vary in their effects on fermentation, tissue and digesta mass accretion, and bacterial populations in the large bowel of pigs. Br. J. Nutr. 2007, 97, 134–144. [Google Scholar] [CrossRef] [Green Version]
- Conlon, M.A.; Kerr, C.A.; McSweeney, C.S.; Dunne, R.A.; Shaw, J.M.; Kang, S.; Bird, A.R.; Morell, M.K.; Lockett, T.J.; Molloy, P.L.; et al. Resistant starches protect against colonic DNA damage and alter microbiota and gene expression in rats fed a western diet. J. Nutr. 2012, 142, 832–840. [Google Scholar] [CrossRef] [Green Version]
- Jha, R.; Rossnagel, B.; Pieper, R.; Van Kessel, A.; Leterme, P. Barley and oat cultivars with diverse carbohydrate composition alter ileal and total tract nutrient digestibility and fermentation metabolites in weaned piglets. Animal 2010, 4, 724–731. [Google Scholar] [CrossRef] [Green Version]
- Le Leu, R.K.; Brown, I.L.; Hu, Y.; Morita, T.; Esterman, A.; Young, G.P. Effect of dietary resistant starch and protein on colonic fermentation and intestinal tumourigenesis in rats. Carcinogenesis 2007, 28, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, F.; Wang, J.; Wang, A.; Yao, X.; Strappe, P.; Zhou, Z.; Wu, Q.; Guo, T. Starch acylation of different short-chain fatty acids and its corresponding influence on gut microbiome and diabetic indexes. Food Chem. 2022, 389, 133089. [Google Scholar] [CrossRef] [PubMed]
- Morita, T.; Kasaoka, S.; Kiriyama, S.; Brown, I.L.; Topping, D.L. Comparative effects of acetylated and unmodified high-amylose maize starch in rats. Starch-Starke 2005, 57, 246–253. [Google Scholar] [CrossRef]
- Nagata, R.; Kamibayashi, R.; Bochimoto, H.; Fukuma, N.; Shimada, K.; Tachibe, M.; Takaishi, Y.; Han, K.-H.; Fukushima, M. Chemical modification of cornstarch by hydroxypropylation enhances cecal fermentation-mediated lipid metabolism in rats. Starch-Starke 2020, 72, 1900050. [Google Scholar] [CrossRef]
- Regmi, P.R.; Metzler-Zebeli, B.U.; Gaenzle, M.G.; van Kempen, T.A.T.G.; Zijlstra, R.T. Starch with high amylose content and low In vitro digestibility increases intestinal nutrient flow and microbial fermentation and selectively promotes bifidobacteria in pigs. J. Nutr. 2011, 141, 1273–1280. [Google Scholar] [CrossRef] [Green Version]
- Saito, K.; Ito, T.; Kuribayashi, T.; Mochida, K.; Nakakuki, T.; Shibata, M.; Sugawara, M. Effect of raw and heat-moisture treated high-amylose corn starch on fermentation by the rat cecal bacteria. Starch-Starke 2001, 53, 424–430. [Google Scholar] [CrossRef]
- Li, M.; Wang, F.; Wang, J.; Wang, R.; Strappe, P.; Zheng, B.; Zhou, Z.; Chen, L. Manipulation of the internal structure of starch by propionyl treatment and its diverse influence on digestion and in vitro fermentation characteristics. Carbohydr. Polym. 2021, 270, 118390. [Google Scholar] [CrossRef]
- Marino, E.; Richards, J.L.; McLeod, K.H.; Stanley, D.; Yap, Y.A.; Knight, J.; McKenzie, C.; Kranich, J.; Oliveira, A.C.; Rossello, F.J.; et al. Gut microbial metabolites limit the frequency of autoimmune T cells and protect against type 1 diabetes. Nat. Immunol. 2017, 18, 552–562. [Google Scholar] [CrossRef]
- Li, H.-T.; Gilbert, R.G.; Gidley, M.J. Molecular-structure evolution during in vitro fermentation of granular high-amylose wheat starch is different to in vitro digestion. Food Chem. 2021, 362, 130188. [Google Scholar] [CrossRef]
- Petropoulou, K.; Salt, L.J.; Edwards, C.H.; Warren, F.J.; Garcia-Perez, I.; Chambers, E.S.; Alshaalan, R.; Khatib, M.; Perez-Moral, N.; Cross, K.L.; et al. A natural mutation in Pisum sativum L. (pea) alters starch assembly and improves glucose homeostasis in humans. Nat. Food 2020, 1, 693–704. [Google Scholar] [CrossRef]
- Gidley, M.J. Give peas a chance. Nat. Food 2020, 1, 663–664. [Google Scholar] [CrossRef]
- Vogelstein, B.; Fearon, E.R.; Hamilton, S.R.; Kern, S.E.; Preisinger, A.C.; Leppert, M.; Nakamura, Y.; White, R.; Smits, A.M.; Bos, J.L. Genetic alterations during colorectal-tumor development. New Engl. J. Med. 1988, 319, 525–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Leu, R.K.; Hu, Y.; Brown, I.L.; Young, G.P. Effect of high amylose maize starches on colonic fermentation and apoptotic response to DNA-damage in the colon of rats. Nutr. Metab. 2009, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gondalia, S.V.; Wymond, B.; Benassi-Evans, B.; Berbezy, P.; Bird, A.R.; Belobrajdic, D.P. Substitution of refined conventional wheat flour with wheat high in resistant starch modulates the intestinal microbiota and fecal metabolites in healthy adults: A randomized, controlled trial. J. Nutr. 2022, 152, 1426–1437. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.; Vuksan, V.; Kendall, C.W.; Würsch, P.; Jeffcoat, R.; Waring, S.; Mehling, C.C.; Vidgen, E.; Augustin, L.S.; Wong, E. Physiological effects of resistant starches on fecal bulk, short chain fatty acids, blood lipids and glycemic index. J. Am. Coll. Nutr. 1998, 17, 609–616. [Google Scholar] [CrossRef]
- Noakes, M.; Clifton, P.M.; Nestel, P.J.; Le Leu, R.; McIntosh, G. Effect of high-amylose starch and oat bran on metabolic variables and bowel function in subjects with hypertriglyceridemia. Am. J. Clin. Nutr. 1996, 64, 944–951. [Google Scholar] [CrossRef] [Green Version]
- Penn-Marshall, M.; Holtzman, G.I.; Barbeau, W.E. African Americans may have to consume more than 12 grams a day of resistant starch to lower their risk for type 2 diabetes. J. Med. Food 2010, 13, 999–1004. [Google Scholar] [CrossRef]
- Sobh, M.; Montroy, J.; Daham, Z.; Sibbald, S.; Lalu, M.; Stintzi, A.; Mack, D.; Fergusson, D.A. Tolerability and SCFA production after resistant starch supplementation in humans: A systematic review of randomized controlled studies. Am. J. Clin. Nutr. 2022, 115, 608–618. [Google Scholar] [CrossRef]
- Anderson, G.H.; Cho, C.E.; Akhavan, T.; Mollard, R.C.; Luhovyy, B.L.; Finocchiaro, E.T. Relation between estimates of cornstarch digestibility by the Englyst in vitro method and glycemic response, subjective appetite, and short-term food intake in young men. Am. J. Clin. Nutr. 2010, 91, 932–939. [Google Scholar] [CrossRef] [Green Version]
- Belobrajdic, D.P.; Regina, A.; Klingner, B.; Zajac, I.; Chapron, S.; Berbezy, P.; Bird, A.R. High-amylose wheat lowers the postprandial glycemic response to bread in healthy adults: A randomized controlled crossover trial. J. Nutr. 2019, 149, 1335–1345. [Google Scholar] [CrossRef] [Green Version]
- Bodinham, C.L.; Smith, L.; Wright, J.; Frost, G.S.; Robertson, M.D. Dietary fibre improves first-phase insulin secretion in overweight individuals. PLoS ONE 2012, 7, e40834. [Google Scholar] [CrossRef] [PubMed]
- Brighenti, F.; Benini, L.; Del Rio, D.; Casiraghi, C.; Pellegrini, N.; Scazzina, F.; Jenkins, D.J.A.; Vantini, I. Colonic fermentation of indigestible carbohydrates contributes to the second-meal effect. Am. J. Clin. Nutr. 2006, 83, 817–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haini, N.; Jau-Shya, L.; Rosli, R.G.M.; Mamat, H. Effects of high-amylose maize starch on the glycemic index of Chinese steamed buns (CSB). Heliyon 2022, 8, e09375. [Google Scholar] [CrossRef] [PubMed]
- Keogh, J.B.; Lau, C.W.H.; Noakes, M.; Bowen, J.; Clifton, P.M. Effects of meals with high soluble fibre, high amylose barley variant on glucose, insulin, satiety and thermic effect of food in healthy lean women. Eur. J. Clin. Nutr. 2007, 61, 597–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luhovyy, B.L.; Mollard, R.C.; Yurchenko, S.; Nunez, M.F.; Berengut, S.; Liu, T.T.; Smith, C.E.; Pelkman, C.L.; Anderson, G.H. The effects of whole grain high-amylose maize flour as a source of resistant starch on blood glucose, satiety, and food intake in young men. J. Food Sci. 2014, 79, H2550–H2556. [Google Scholar] [CrossRef] [PubMed]
- Maziarz, M.P.; Preisendanz, S.; Juma, S.; Imrhan, V.; Prasad, C.; Vijayagopal, P. Resistant starch lowers postprandial glucose and leptin in overweight adults consuming a moderate-to-high-fat diet: A randomized-controlled trial. Nutr. J. 2017, 16, 14. [Google Scholar] [CrossRef] [Green Version]
- Stewart, M.L.; Zimmer, J.P. Postprandial glucose and insulin response to a high-fiber muffin top containing resistant starch type 4 in healthy adults: A double-blind, randomized, controlled trial. Nutrition 2018, 53, 59–63. [Google Scholar] [CrossRef]
- Robertson, M.D.; Wright, J.W.; Loizon, E.; Debard, C.; Vidal, H.; Shojaee-Moradie, F.; Russell-Jones, D.; Umpleby, A.M. Insulin-sensitizing effects on muscle and adipose tissue after dietary fiber intake in men and women with metabolic syndrome. J. Clin. Endocrinol. Metab. 2012, 97, 3326–3332. [Google Scholar] [CrossRef] [Green Version]
- Ames, N.; Blewett, H.; Storsley, J.; Thandapilly, S.J.; Zahradka, P.; Taylor, C. A double-blind randomised controlled trial testing the effect of a barley product containing varying amounts and types of fibre on the postprandial glucose response of healthy volunteers. Br. J. Nutr. 2015, 113, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Maki, K.C.; Pelkman, C.L.; Finocchiaro, E.T.; Kelley, K.M.; Lawless, A.L.; Schild, A.L.; Rains, T.M. Resistant starch from high-amylose maize increases insulin sensitivity in overweight and obese men. J. Nutr. 2012, 142, 717–723. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, A.C.; Ostman, E.M.; Hoist, J.J.; Bjorck, I.M.E. Including indigestible carbohydrates in the evening meal of healthy subjects improves glucose tolerance, lowers inflammatory markers, and increases satiety after a subsequent standardized breakfast. J. Nutr. 2008, 138, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Al-Mana, N.M.; Robertson, M.D. Acute effect of resistant starch on food intake, appetite and satiety in overweight/obese males. Nutrients 2018, 10, 1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gower, B.A.; Bergman, R.; Stefanovski, D.; Darnell, B.; Ovalle, F.; Fisher, G.; Sweatt, S.K.; Resuehr, H.S.; Pelkman, C. Baseline insulin sensitivity affects response to high-amylose maize resistant starch in women: A randomized, controlled trial. Nutr. Metab. 2016, 13, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dainty, S.A.; Klingel, S.L.; Pilkey, S.E.; McDonald, E.; McKeown, B.; Emes, M.J.; Duncan, A.M. Resistant starch bagels reduce fasting and postprandial insulin in adults at risk of type 2 diabetes. J. Nutr. 2016, 146, 2252–2259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, K.F. An introductory review of resistant starch type 2 from high-amylose cereal grains and its effect on glucose and insulin homeostasis. Nutr. Rev. 2019, 77, 748–764. [Google Scholar] [CrossRef] [Green Version]
- Robertson, M.D.; Bickerton, A.S.; Vidal, H.; Frayn, K.N. Insulin-sensitizing effects of dietary resistant starch and effects on skeletal muscle and adipose tissue metabolism. Am. J. Clin. Nutr. 2005, 82, 559–567. [Google Scholar] [CrossRef]
- Firth, J.; Gangwisch, J.E.; Borisini, A.; Wootton, R.E.; Mayer, E.A. Food and mood: How do diet and nutrition affect mental wellbeing? BMJ 2020, 369, m2382. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.-T.; Zhang, W.; Zhu, H.; Chao, C.; Guo, Q. Unlocking the Potential of High-Amylose Starch for Gut Health: Not All Function the Same. Fermentation 2023, 9, 134. https://doi.org/10.3390/fermentation9020134
Li H-T, Zhang W, Zhu H, Chao C, Guo Q. Unlocking the Potential of High-Amylose Starch for Gut Health: Not All Function the Same. Fermentation. 2023; 9(2):134. https://doi.org/10.3390/fermentation9020134
Chicago/Turabian StyleLi, Hai-Teng, Wenyu Zhang, Huijuan Zhu, Chen Chao, and Qing Guo. 2023. "Unlocking the Potential of High-Amylose Starch for Gut Health: Not All Function the Same" Fermentation 9, no. 2: 134. https://doi.org/10.3390/fermentation9020134
APA StyleLi, H. -T., Zhang, W., Zhu, H., Chao, C., & Guo, Q. (2023). Unlocking the Potential of High-Amylose Starch for Gut Health: Not All Function the Same. Fermentation, 9(2), 134. https://doi.org/10.3390/fermentation9020134