Exploiting the Fruitfly, Drosophila melanogaster, to Identify the Molecular Basis of Cryptochrome-Dependent Magnetosensitivity


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Main Text
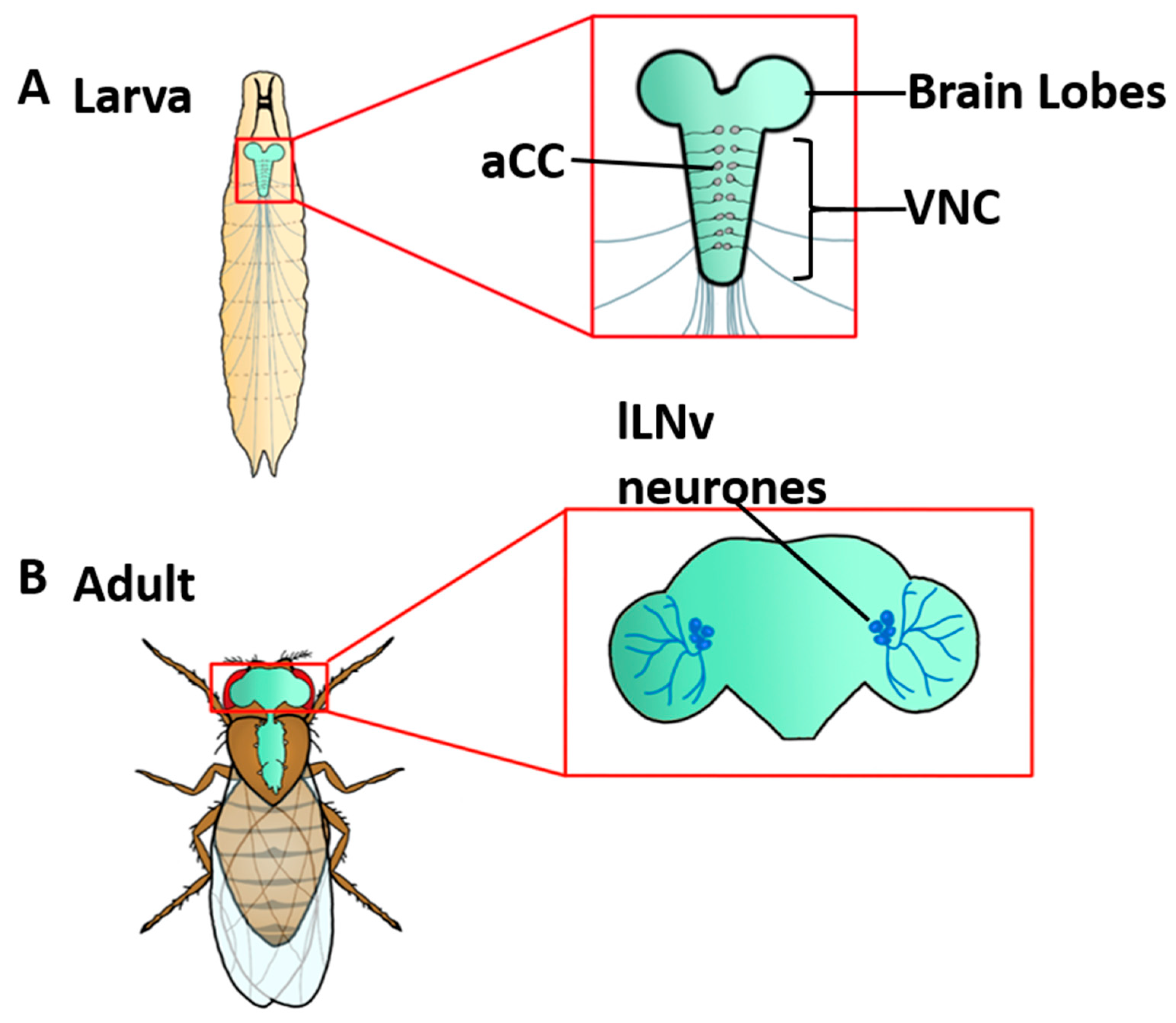
2. Drosophila Magnetosensitivity Requires the Presence of Cryptochrome
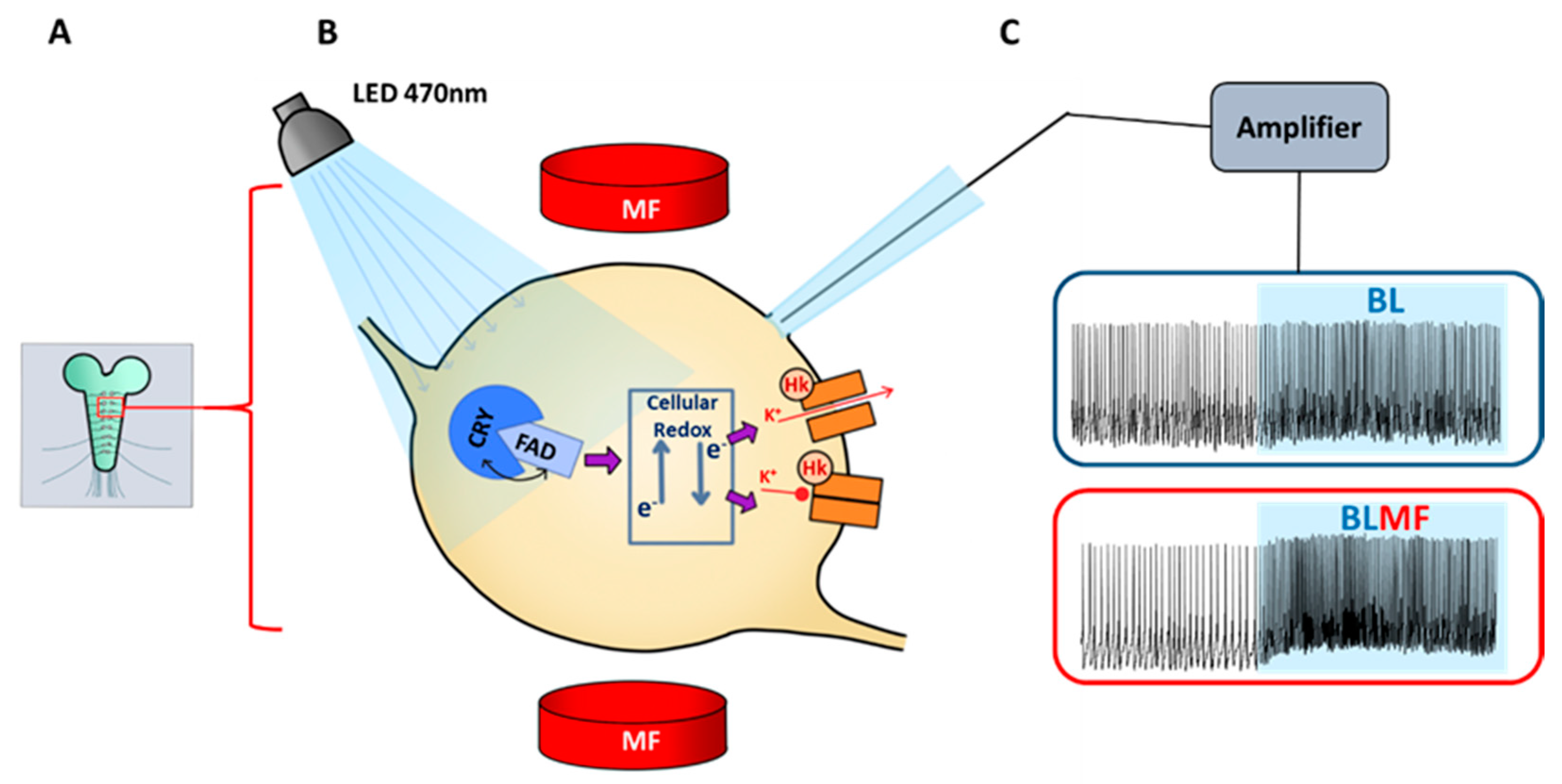
3. Mechanistic Basis of Magnetosensitivity in Drosophila
4. CRY Mediates Magnetosensitivity in Drosophila Neurons
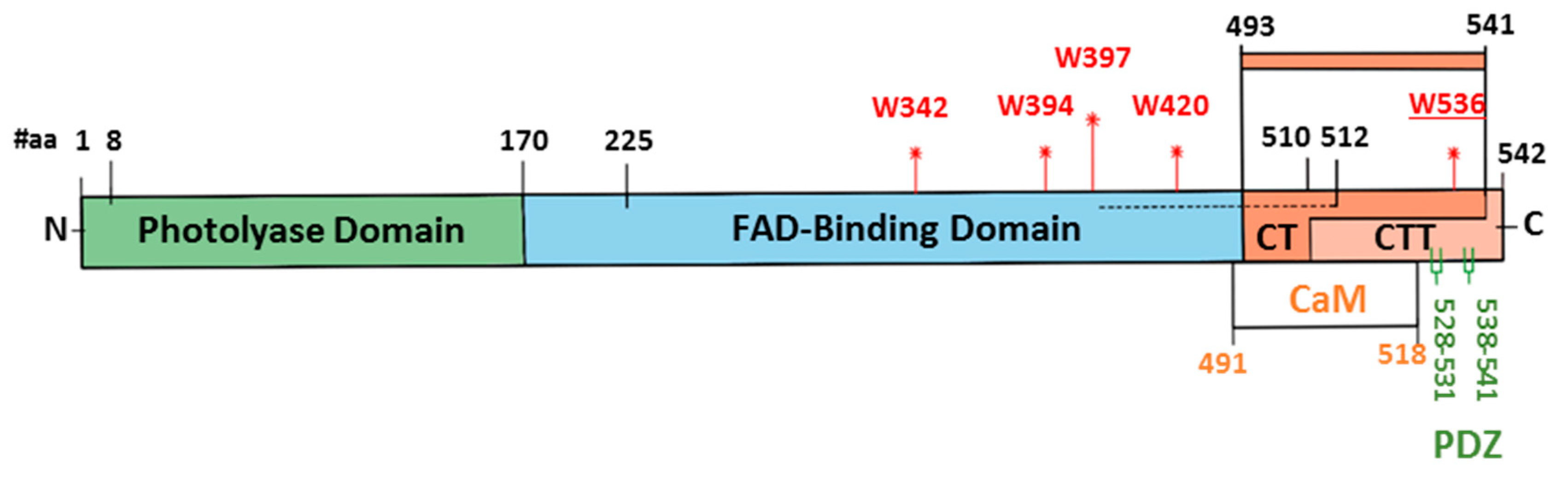
5. Is Full-Length CRY Essential for Magnetosensitivity in Drosophila?
6. Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schulten, K.; Staerk, H.; Weller, A.; Werner, H.J.; Nickel, B. Magnetic field dependence of the geminate recombination of radical ion pairs in polar solvents. Z. Phys. Chem. 1976, 101, 371–390. [Google Scholar] [CrossRef]
- Ritz, T.; Adem, S.; Schulten, K. A model for photoreceptor-based magnetoreception in birds. Biophys. J. 2000, 78, 707–718. [Google Scholar] [CrossRef] [Green Version]
- Hore, P.J.; Mouritsen, H. The Radical-Pair Mechanism of Magnetoreception. Annu. Rev. Biophys. 2016, 45, 299–344. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, A.T.; Top, D.; Manahan, C.C.; Tokuda, J.M.; Zhang, S.; Pollack, L.; Young, M.W.; Crane, B.R. Flavin reduction activates Drosophila cryptochrome. Proc. Natl. Acad. Sci. USA 2013, 110, 20455–20460. [Google Scholar] [CrossRef] [Green Version]
- Vacha, M. Magnetoreception of Invertebrates; Oxford University Press: Oxford, UK, 2019. [Google Scholar]
- Damulewicz, M.; Mazzotta, G.M. One Actor, Multiple Roles: The Performances of Cryptochrome in Drosophila. Front. Physiol. 2020, 11, 99. [Google Scholar] [CrossRef] [Green Version]
- Foley, L.E.; Emery, P. Drosophila Cryptochrome: Variations in Blue. J. Biol. Rhythms. 2020, 35, 16–27. [Google Scholar] [CrossRef]
- Wehner, R.; Labhart, T. Perception of the geomagnetic field in the fly Drosophila melanogaster. Experientia 1970, 26, 967–968. [Google Scholar] [CrossRef]
- Phillips, J.B.; Sayeed, O. Wavelength-dependent effects of light on magnetic compass orientation in Drosophila melanogaster. J. Comp. Physiol. A 1993, 172, 303–308. [Google Scholar] [CrossRef]
- Gegear, R.J.; Casselman, A.; Waddell, S.; Reppert, S.M. Cryptochrome mediates light-dependent magnetosensitivity in Drosophila. Nature 2008, 454, 1014–1018. [Google Scholar] [CrossRef] [Green Version]
- Gegear, R.J.; Foley, L.E.; Casselman, A.; Reppert, S.M. Animal cryptochromes mediate magnetoreception by an unconventional photochemical mechanism. Nature 2010, 463, 804–807. [Google Scholar] [CrossRef]
- Foley, L.E.; Gegear, R.J.; Reppert, S.M. Human cryptochrome exhibits light-dependent magnetosensitivity. Nat. Commun. 2011, 2, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshii, T.; Ahmad, M.; Helfrich-Forster, C. Cryptochrome mediates light-dependent magnetosensitivity of Drosophila’s circadian clock. PLoS Biol. 2009, 7, e1000086. [Google Scholar] [CrossRef] [PubMed]
- Fedele, G.; Edwards, M.D.; Bhutani, S.; Hares, J.M.; Murbach, M.; Green, E.W.; Dissel, S.; Hastings, M.H.; Rosato, E.; Kyriacou, C.P. Genetic analysis of circadian responses to low frequency electromagnetic fields in Drosophila melanogaster. PLoS Genet. 2014, 10, e1004804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, J.B.; Jorge, P.E.; Muheim, R. Light-dependent magnetic compass orientation in amphibians and insects: Candidate receptors and candidate molecular mechanisms. J. R. Soc. Interface 2010, 7 (Suppl. S2), S241–S256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedele, G.; Green, E.W.; Rosato, E.; Kyriacou, C.P. An electromagnetic field disrupts negative geotaxis in Drosophila via a CRY-dependent pathway. Nat. Commun. 2014, 5, 4391. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.E.; Bang, S.; Min, S.; Lee, S.H.; Kwon, S.H.; Lee, Y.; Lee, Y.H.; Chung, J.; Chae, K.S. Positive geotactic behaviors induced by geomagnetic field in Drosophila. Mol. Brain 2016, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.H.; Chuang, C.L.; Jiang, J.A.; Yang, E.C. Magnetic Sensing through the Abdomen of the Honey bee. Sci Rep. 2016, 6, 23657. [Google Scholar] [CrossRef] [Green Version]
- Bazalova, O.; Kvicalova, M.; Valkova, T.; Slaby, P.; Bartos, P.; Netusil, R.; Tomanova, K.; Braeunig, P.; Lee, H.J.; Sauman, I.; et al. Cryptochrome 2 mediates directional magnetoreception in cockroaches. Proc. Natl. Acad. Sci. USA 2016, 113, 1660–1665. [Google Scholar] [CrossRef] [Green Version]
- Slaby, P.; Bartos, P.; Karas, J.; Netusil, R.; Tomanova, K.; Vacha, M. How Swift Is Cry-Mediated Magnetoreception? Conditioning in an American Cockroach Shows Sub-second Response. Front. Behav. Neurosci. 2018, 12, 107. [Google Scholar] [CrossRef] [Green Version]
- Wiltschko, W.; Wiltschko, R. Magnetic orientation and magnetoreception in birds and other animals. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2005, 191, 675–693. [Google Scholar] [CrossRef]
- Johnsen, S.; Mattern, E.; Ritz, T. Light-dependent magnetoreception: Quantum catches and opponency mechanisms of possible photosensitive molecules. J. Exp. Biol. 2007, 210, 3171–3178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.; Robertson, D.E.; Ahmad, M.; Raibekas, A.A.; Jorns, M.S.; Dutton, P.L.; Cashmore, A.R. Association of flavin adenine dinucleotide with the Arabidopsis blue light receptor CRY1. Science 1995, 269, 968–970. [Google Scholar] [CrossRef]
- Berndt, A.; Kottke, T.; Breitkreuz, H.; Dvorsky, R.; Hennig, S.; Alexander, M.; Wolf, E. A novel photoreaction mechanism for the circadian blue light photoreceptor Drosophila cryptochrome. J. Biol. Chem. 2007, 282, 13011–13021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouly, J.P.; Schleicher, E.; Dionisio-Sese, M.; Vandenbussche, F.; Van Der Straeten, D.; Bakrim, N.; Meier, S.; Batschauer, A.; Galland, P.; Bittl, R.; et al. Cryptochrome blue light photoreceptors are activated through interconversion of flavin redox states. J. Biol. Chem. 2007, 282, 9383–9391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, N.; Bouly, J.P.; Ahmad, M. Evidence of a light-sensing role for folate in Arabidopsis cryptochrome blue-light receptors. Mol. Plant. 2008, 1, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, R.; Schleicher, E.; Meier, S.; Viana, R.M.; Pokorny, R.; Ahmad, M.; Bittl, R.; Batschauer, A. The signaling state of Arabidopsis cryptochrome 2 contains flavin semiquinone. J. Biol. Chem. 2007, 282, 14916–14922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dissel, S.; Hansen, C.N.; Ozkaya, O.; Hemsley, M.; Kyriacou, C.P.; Rosato, E. The logic of circadian organization in Drosophila. Curr. Biol. 2014, 24, 2257–2266. [Google Scholar] [CrossRef] [Green Version]
- Busza, A.; Emery-Le, M.; Rosbash, M.; Emery, P. Roles of the two Drosophila CRYPTOCHROME structural domains in circadian photoreception. Science 2004, 304, 1503–1506. [Google Scholar] [CrossRef] [Green Version]
- Peschel, N.; Chen, K.F.; Szabo, G.; Stanewsky, R. Light-dependent interactions between the Drosophila circadian clock factors cryptochrome, jetlag, and timeless. Curr. Biol. 2009, 19, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Hemsley, M.J.; Mazzotta, G.M.; Mason, M.; Dissel, S.; Toppo, S.; Pagano, M.A.; Sandrelli, F.; Meggio, F.; Rosato, E.; Costa, R.; et al. Linear motifs in the C-terminus of D. melanogaster cryptochrome. Biochem. Biophys. Res. Commun. 2007, 355, 531–537. [Google Scholar] [CrossRef]
- Mazzotta, G.M.; Bellanda, M.; Minervini, G.; Damulewicz, M.; Cusumano, P.; Aufiero, S.; Stefani, M.; Zambelli, B.; Mammi, S.; Costa, R.; et al. Calmodulin Enhances Cryptochrome Binding to INAD in Drosophila Photoreceptors. Front. Mol. Neurosci. 2018, 11, 280. [Google Scholar] [CrossRef] [PubMed]
- Muller, P.; Ahmad, M. Light-activated cryptochrome reacts with molecular oxygen to form a flavin-superoxide radical pair consistent with magnetoreception. J. Biol. Chem. 2011, 286, 21033–21040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consentino, L.; Lambert, S.; Martino, C.; Jourdan, N.; Bouchet, P.E.; Witczak, J.; Castello, P.; El-Esawi, M.; Corbineau, F.; d′Harlingue, A.; et al. Blue-light dependent reactive oxygen species formation by Arabidopsis cryptochrome may define a novel evolutionarily conserved signaling mechanism. New Phytol. 2015, 206, 1450–1462. [Google Scholar] [CrossRef] [PubMed]
- Fogle, K.J.; Baik, L.S.; Houl, J.H.; Tran, T.T.; Roberts, L.; Dahm, N.A.; Cao, Y.; Zhou, M.; Holmes, T.C. CRYPTOCHROME-mediated phototransduction by modulation of the potassium ion channel beta-subunit redox sensor. Proc. Natl. Acad. Sci. USA 2015, 112, 2245–2250. [Google Scholar] [CrossRef] [Green Version]
- Pooam, M.; Arthaut, L.D.; Burdick, D.; Link, J.; Martino, C.F.; Ahmad, M. Magnetic sensitivity mediated by the Arabidopsis blue-light receptor cryptochrome occurs during flavin reoxidation in the dark. Planta 2019, 249, 319–332. [Google Scholar] [CrossRef] [Green Version]
- El-Esawi, M.; Arthaut, L.D.; Jourdan, N.; d′Harlingue, A.; Link, J.; Martino, C.F.; Ahmad, M. Blue-light induced biosynthesis of ROS contributes to the signaling mechanism of Arabidopsis cryptochrome. Sci. Rep. 2017, 7, 13875. [Google Scholar] [CrossRef]
- Arthaut, L.D.; Jourdan, N.; Mteyrek, A.; Procopio, M.; El-Esawi, M.; d′Harlingue, A.; Bouchet, P.E.; Witczak, J.; Ritz, T.; Klarsfeld, A.; et al. Blue-light induced accumulation of reactive oxygen species is a consequence of the Drosophila cryptochrome photocycle. PLoS ONE 2017, 12, e0171836. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Robinson, A.J.; Henbest, K.B.; Hogben, H.J.; Biskup, T.; Ahmad, M.; Schleicher, E.; Weber, S.; Timmel, C.R.; Hore, P.J. Magnetically sensitive light-induced reactions in cryptochrome are consistent with its proposed role as a magnetoreceptor. Proc. Natl. Acad. Sci. USA 2012, 109, 4774–4779. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, D.M.; Li, J.; Henbest, K.B.; Neil, S.R.; Maeda, K.; Storey, J.; Schleicher, E.; Biskup, T.; Rodriguez, R.; Weber, S.; et al. Millitesla magnetic field effects on the photocycle of an animal cryptochrome. Sci. Rep. 2017, 7, 42228. [Google Scholar] [CrossRef] [Green Version]
- Fogle, K.J.; Parson, K.G.; Dahm, N.A.; Holmes, T.C. CRYPTOCHROME is a blue-light sensor that regulates neuronal firing rate. Science 2011, 331, 1409–1413. [Google Scholar] [CrossRef] [Green Version]
- Buhl, E.; Bradlaugh, A.; Ogueta, M.; Chen, K.F.; Stanewsky, R.; Hodge, J.J. Quasimodo mediates daily and acute light effects on Drosophila clock neuron excitability. Proc. Natl. Acad. Sci. USA 2016, 113, 13486–13491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giachello, C.N.G.; Scrutton, N.S.; Jones, A.R.; Baines, R.A. Magnetic Fields Modulate Blue-Light-Dependent Regulation of Neuronal Firing by Cryptochrome. J. Neurosci. 2016, 36, 10742–10749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chouinard, S.W.; Wilson, G.F.; Schlimgen, A.K.; Ganetzky, B. A potassium channel beta subunit related to the aldo-keto reductase superfamily is encoded by the Drosophila hyperkinetic locus. Proc. Natl. Acad. Sci. USA 1995, 92, 6763–6767. [Google Scholar] [CrossRef] [Green Version]
- Kempf, A.; Song, S.M.; Talbot, C.B.; Miesenbock, G. A potassium channel beta-subunit couples mitochondrial electron transport to sleep. Nature 2019, 568, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Marley, R.; Giachello, C.N.; Scrutton, N.S.; Baines, R.A.; Jones, A.R. Cryptochrome-dependent magnetic field effect on seizure response in Drosophila larvae. Sci. Rep. 2014, 4, 5799. [Google Scholar] [CrossRef] [Green Version]
- Steiner, U.E.; Ulrich, T. Magnetic field effects in chemical kinetics and related phenomena. Chem. Rev. 1989, 89, 51–147. [Google Scholar] [CrossRef] [Green Version]
- Muller, P.; Brettel, K.; Grama, L.; Nyitrai, M.; Lukacs, A. Photochemistry of Wild-Type and N378D Mutant E. coli DNA Photolyase with Oxidized FAD Cofactor Studied by Transient Absorption Spectroscopy. Chemphyschem 2016, 17, 1329–1340. [Google Scholar] [CrossRef]
- Nohr, D.; Franz, S.; Rodriguez, R.; Paulus, B.; Essen, L.O.; Weber, S.; Schleicher, E. Extended Electron-Transfer in Animal Cryptochromes Mediated by a Tetrad of Aromatic Amino Acids. Biophys. J. 2016, 111, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Top, D.; Manahan, C.C.; Young, M.W.; Crane, B.R. Circadian clock activity of cryptochrome relies on tryptophan-mediated photoreduction. Proc. Natl. Acad. Sci. USA 2018, 115, 3822–3827. [Google Scholar] [CrossRef] [Green Version]
- Baik, L.S.; Au, D.D.; Nave, C.; Foden, A.J.; Enrriquez-Villalva, W.K.; Holmes, T.C. Distinct mechanisms of Drosophila CRYPTOCHROME-mediated light-evoked membrane depolarization and in vivo clock resetting. Proc. Natl. Acad. Sci. USA 2019, 116, 23339–23344. [Google Scholar] [CrossRef] [Green Version]
- Solov′yov, I.A.; Schulten, K. Magnetoreception through cryptochrome may involve superoxide. Biophys. J. 2009, 96, 4804–4813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiltschko, R.; Ahmad, M.; Niessner, C.; Gehring, D.; Wiltschko, W. Light-dependent magnetoreception in birds: The crucial step occurs in the dark. J. R. Soc. Interface 2016, 13, 20151010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutta, R.J.; Archipowa, N.; Johannissen, L.O.; Jones, A.R.; Scrutton, N.S. Vertebrate Cryptochromes are Vestigial Flavoproteins. Sci. Rep. 2017, 7, 44906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antill, L.M.; Woodward, J.R. Flavin Adenine Dinucleotide Photochemistry Is Magnetic Field Sensitive at Physiological pH. J. Phys. Chem. Lett. 2018, 9, 2691–2696. [Google Scholar] [CrossRef]
- Ikeya, N.; Woodward, J.R. Cellular autofluorescence is magnetic field sensitive. Proc. Natl. Acad. Sci. USA 2021, 118, e2018043118. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bradlaugh, A.; Munro, A.L.; Jones, A.R.; Baines, R.A. Exploiting the Fruitfly, Drosophila melanogaster, to Identify the Molecular Basis of Cryptochrome-Dependent Magnetosensitivity. Quantum Rep. 2021, 3, 127-136. https://doi.org/10.3390/quantum3010007
Bradlaugh A, Munro AL, Jones AR, Baines RA. Exploiting the Fruitfly, Drosophila melanogaster, to Identify the Molecular Basis of Cryptochrome-Dependent Magnetosensitivity. Quantum Reports. 2021; 3(1):127-136. https://doi.org/10.3390/quantum3010007
Chicago/Turabian StyleBradlaugh, Adam, Anna L. Munro, Alex R. Jones, and Richard A. Baines. 2021. "Exploiting the Fruitfly, Drosophila melanogaster, to Identify the Molecular Basis of Cryptochrome-Dependent Magnetosensitivity" Quantum Reports 3, no. 1: 127-136. https://doi.org/10.3390/quantum3010007
APA StyleBradlaugh, A., Munro, A. L., Jones, A. R., & Baines, R. A. (2021). Exploiting the Fruitfly, Drosophila melanogaster, to Identify the Molecular Basis of Cryptochrome-Dependent Magnetosensitivity. Quantum Reports, 3(1), 127-136. https://doi.org/10.3390/quantum3010007