
Reducing Enteric Methanogenesis through Alternate Hydrogen Sinks in the Rumen


Abstract
:1. Introduction
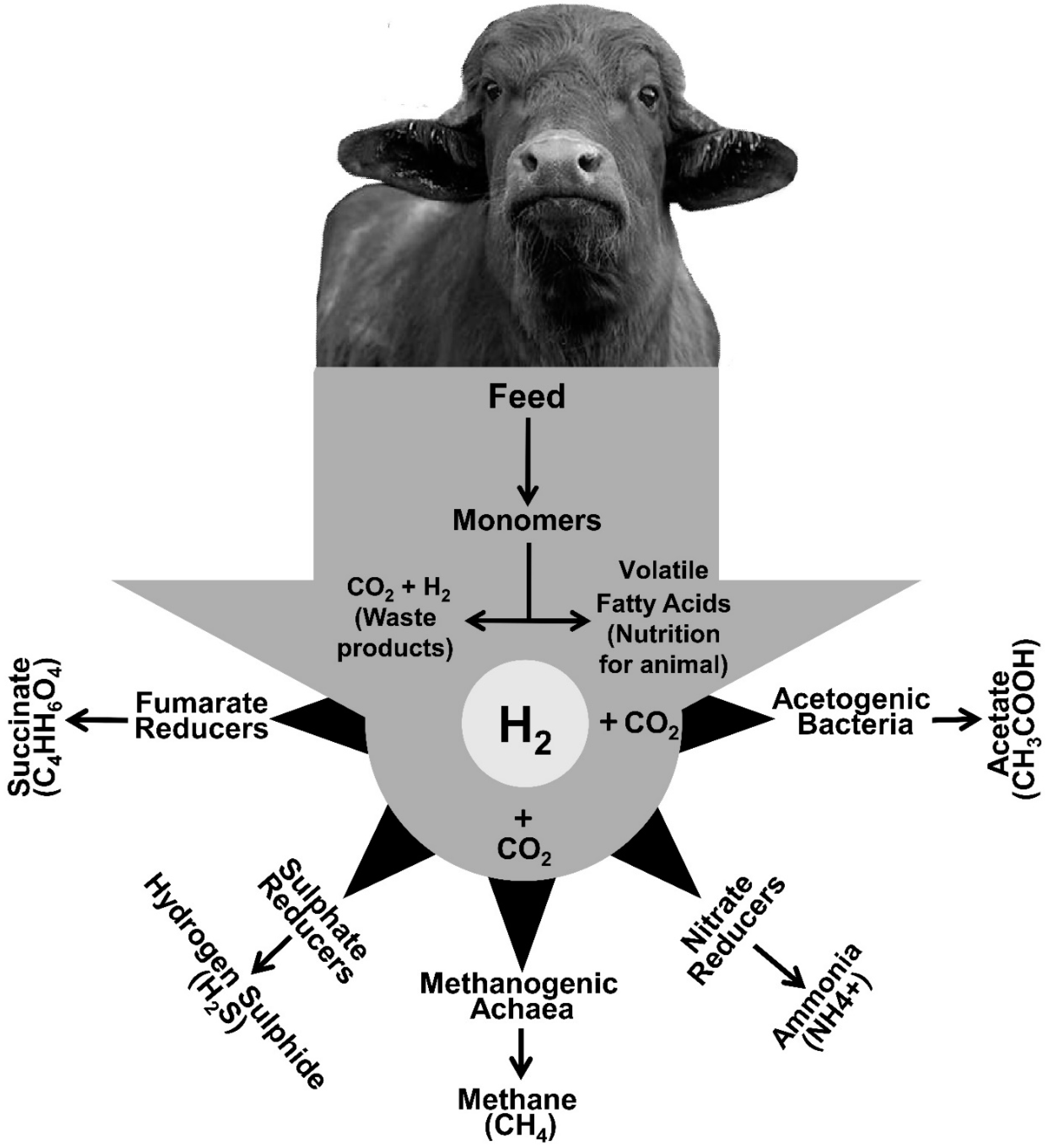
2. Hydrogen: A Key Player in Rumen Fermentation
3. Alternative H2 Sinks
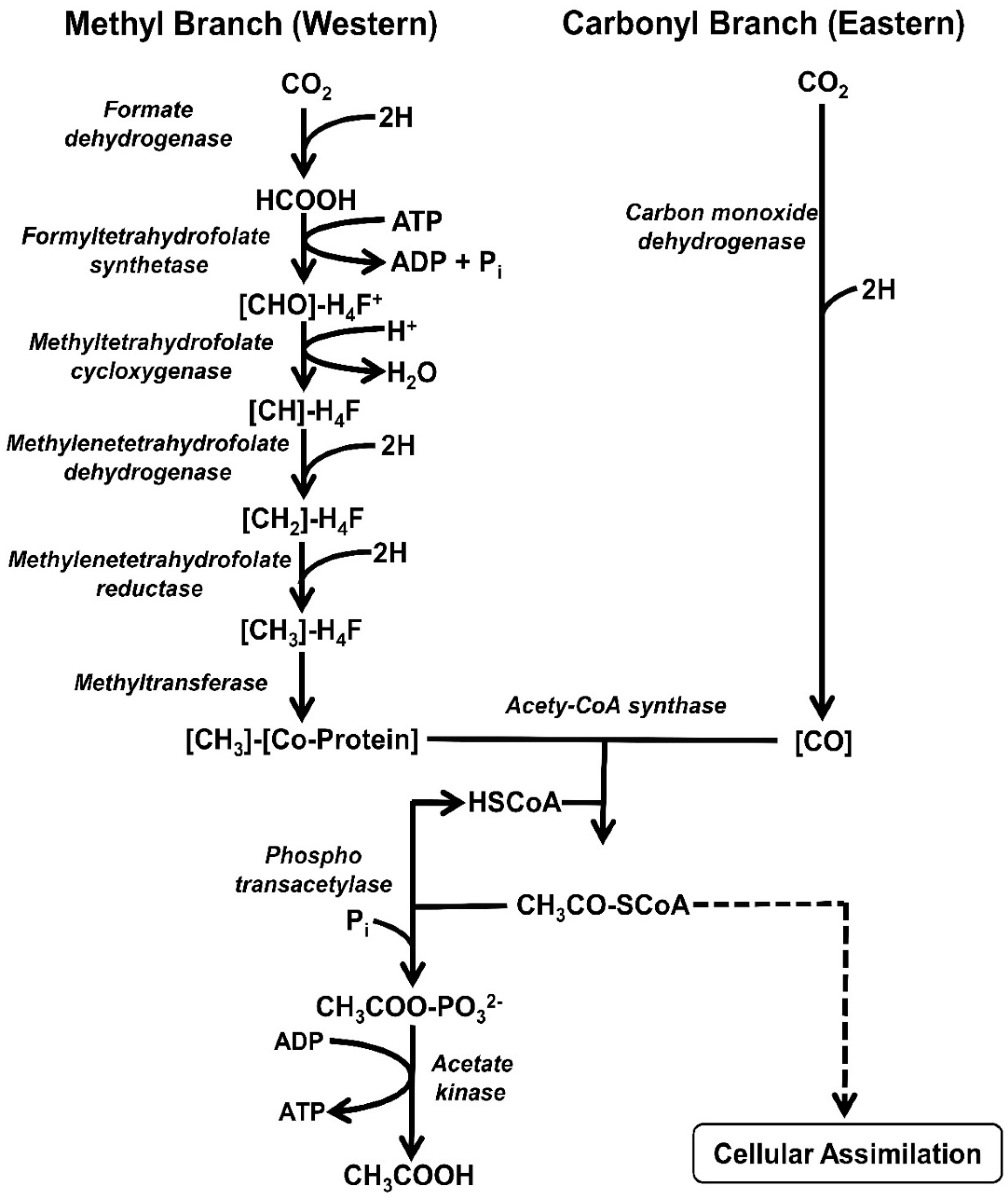
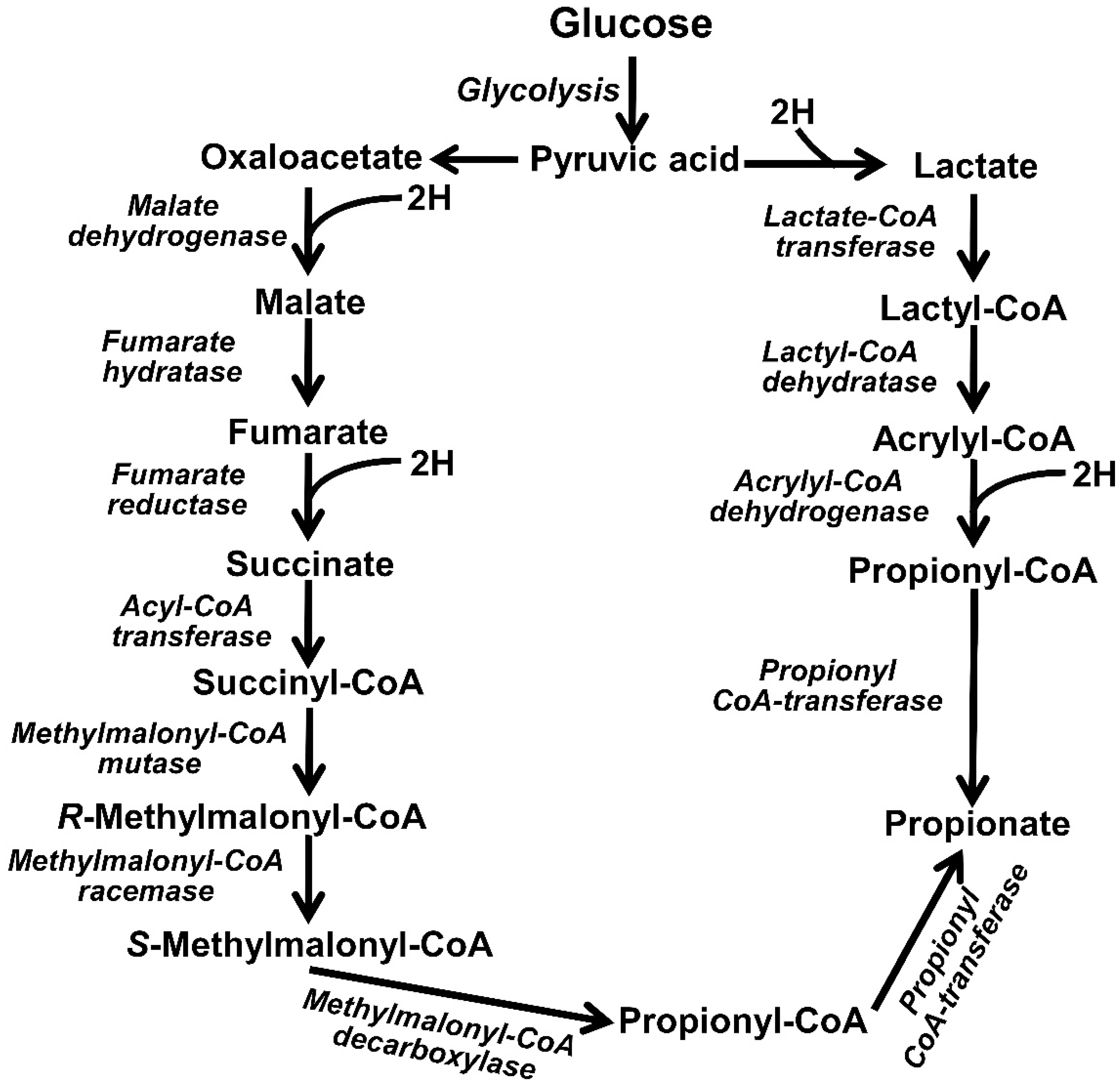
3.1. Reductive Acetogenesis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Categories | Sub Groups | End Products | Microbes (Examples) | Overall Reaction | ΔG0 (kJ) | Problems Associated | Management Strategies | Reference(s) |
|---|---|---|---|---|---|---|---|---|
| Methanogenic sinks | Methnogenesis | Methane (CH4) | Methanobrevibacter ruminantium, Methanomicrobium mobile, Methanobacterium bryantii, Methanobrevibacter smithii, Methanosarcina barkeri, Methanoculleus olentangyi | 4H2 + CO2 → CH4 + 2H2O | −134.0 | Source of ruminal CH4, but not desirable as potent GHG. | Releases H2 accumulation in rumen and need to be suppressed. | [2,4,5,6,11,12,18,20,22,23,24,25,45,63] |
| Non-Methano-genic Sinks | Sulfate Reduction | Hydrogen Sulfide (H2S) | Desulfovibrio desulfuricans, D. vulgaris, Desulfatomaculum spp. | 4H2 + 2H+ + SO4 → H2S + 4 H2O | −234.0 | Undesirable reaction in rumen owing to toxicity of H2S. | Most energy efficient sink in rumen dietary level and feeding strategy must taken into account. | [53,55,70,71,72,73,74,75] |
| Reductive acetogens | Acetic acid (CH3-COOH) | Eubacterium limosum, Acetitomaculum ruminis, Blautia spp, Clostridium spp., Peptostreptococcus productus, Ruminococcus schinkii, Clostridium difficile | 4H2 + 2CO2 → CH3COO− + H+ + 2H2O | −71.6 | Desirable, but needs high levels of H2 partial pressure. | Alteration of rumen microflora with a low H2 threshold possessing capacity for reductive acetogenesis. | [7,10,19,47,54,66,76,77] | |
| Nitrate Reduction | Ammonia (NH4) | Selenomonas ruminantium, Veillonella parvula and Wolinella succinogenes | 4H2 + 2H+ + NO3− → NH4+ + 3H2O | −519.0 | Undesirable reaction in rumen owing to possible accumulation of toxic nitrite. | Gradual adaption of animal to supplement used and development of favorable microflora. | [53,67,70,78,79,80,81,82,83,84,85,86,87,88,89,90,91] | |
| Propionogenesis | Propionic acid (CH3CH2COOH) | Fibrobacter succinogenes, Selenomonas ruminantium ssp. ruminantium, Selenomonas ruminantium ssp. lactilytica, Veillonella parvula and Wolinella succinogenes | C6H12O6 + 2H2→ 2CH3CH2COOH + 2H2O | −84.0 (Fumarate to succinate) | Desirable reaction, but required substrate is costly. | Balancing minimum level in diet and dosing desired microbes governing propionate synthesis. | [57,92,93,94,95,96,97,98,99,100,101,102,103] |
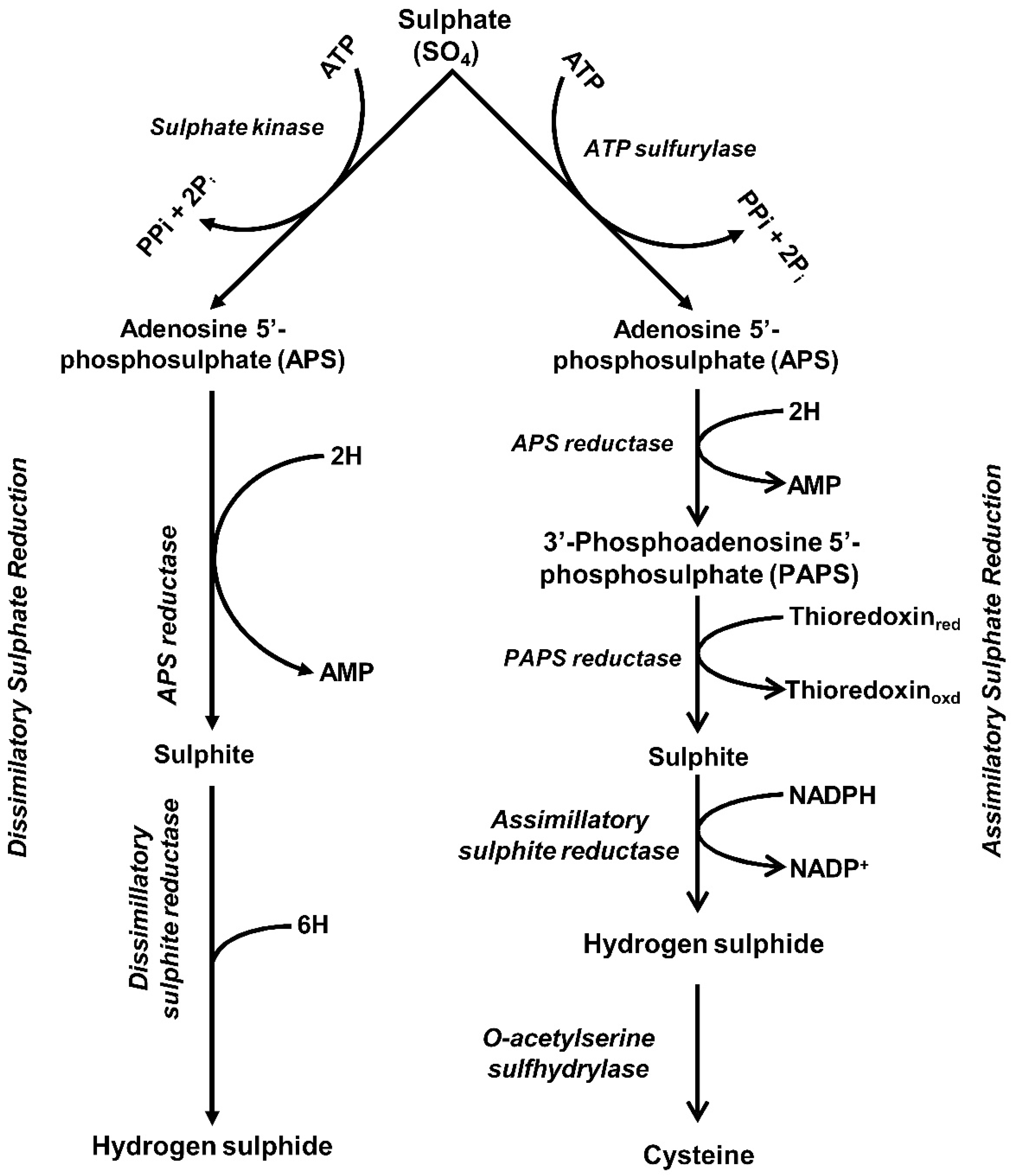
3.2. Sulfur Reduction
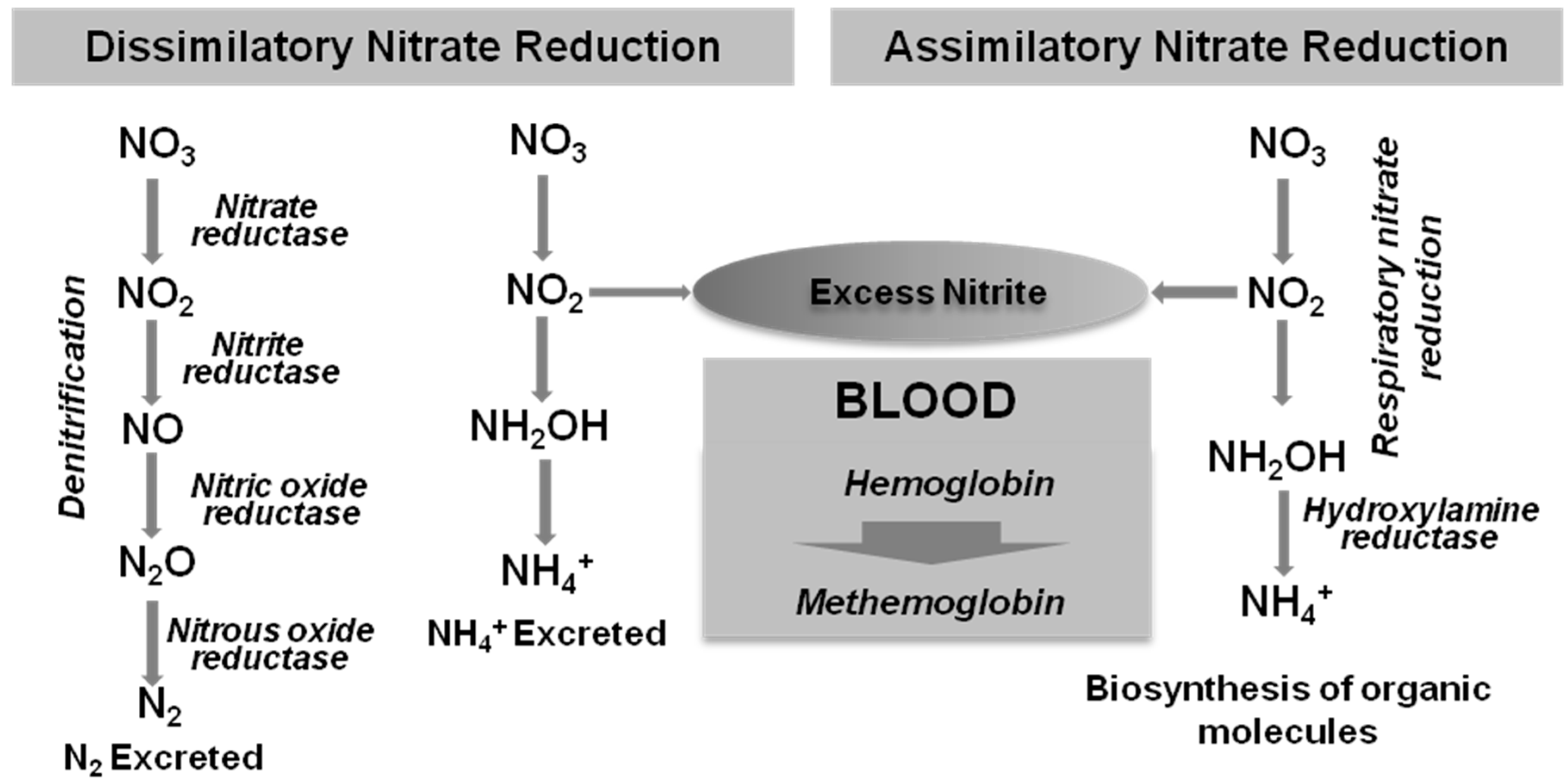
3.3. Nitrate Reduction
3.4. Propionogenesis
4. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, S.; Giller, K.; Kreuzer, M.; Ulbrich, S.E.; Braun, U.; Schwarm, A. Contribution of ruminal fungi, archaea, protozoa, and bacteria to the methane suppression caused by oilseed supplemented diets. Front. Microbiol. 2017, 8, 1864. [Google Scholar] [CrossRef]
- Choudhury, P.K.; Salem, A.Z.M.; Jena, R.; Kumar, S.; Singh, R.; Puniya, A.K. Rumen microbiology: An overview. In Rumen Microbiology-Evolution to Revolution; Puniya, A.K., Singh, R., Kamra, D.N., Eds.; CRC Springer: New Delhi, India, 2015; pp. 3–16. [Google Scholar]
- Kumar, S.; Dagar, S.S.; Puniya, A.K.; Upadhyay, R.C. Changes in methane emission, rumen fermentation in response to diet and microbial interactions. Res. Vet. Sci. 2013, 94, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Kamra, D.N. Rumen microbial ecosystem. Curr. Sci. 2005, 89, 124–135. [Google Scholar]
- Janssen, P.H.; Kirs, M. Structure of the archaeal community of the rumen. Appl. Environ. Microbiol. 2008, 74, 3619–3625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Choudhury, P.K.; Carro, M.D.; Griffith, G.W.; Dagar, S.S.; Puniya, M.; Calabro, S.; Ravella, S.R.; Dhewa, T.; Upadhyay, R.C.; et al. New aspects and strategies for methane mitigation from ruminants. Appl. Microbiol. Biotechnol. 2014, 98, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Ragsdale, S.W.; Pierce, E. Acetogenesis and the Wood-Ljungdahl pathway of CO(2) fixation. Biochim. Biophys. Acta 2008, 1784, 1873–1898. [Google Scholar] [CrossRef] [Green Version]
- Ishaq, S.L.; AlZahal, O.; Walker, N.; McBride, B. An investigation into rumen fungal and protozoal diversity in three rumen fractions, during high-fiber or grain-induced sub-acute ruminal acidosis conditions, with or without active dry yeast supplementation. Front. Microbiol. 2017, 8, 1943. [Google Scholar] [CrossRef] [Green Version]
- Malik, P.K.; Bhatta, R.; Gagen, E.J.; Sejian, V.; Soren, N.M.; Prasad, C.S. Alternate H2 sinks for reducing rumen methanogenesis. In Climate Change Impact on Livestock: Adaptation and Mitigation; Sejian, V., Gaughan, J., Baumgard, L., Prasad, C., Eds.; Springer: New Delhi, India, 2015; pp. 303–320. [Google Scholar]
- Joblin, K. Methanogenic archaea. In Methods in Gut Microbial Ecology for Ruminants; Makker, H., McSweeney, C.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 47–53. [Google Scholar]
- Moss, A.R.; Jouany, J.P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. Ann. Zootech. 2000, 49, 231–253. [Google Scholar] [CrossRef] [Green Version]
- Ungerfeld, E.M. Inhibition of rumen methanogenesis and ruminant productivity: A meta-analysis. Front. Vet. Sci. 2018, 5, 113. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty years of research on rumen methanogenesis: Lessons learned and future challenges for mitigation. Animal 2020, 14, s2–s16. [Google Scholar] [CrossRef] [Green Version]
- Van Nevel, C.J.; Demeyer, D.I. Control of rumen methanogenesis. Environ. Monit. Assess. 1996, 42, 73–97. [Google Scholar] [CrossRef] [PubMed]
- Demeyer, D.; Fiedler, D.; De Graeve, K.G. Attempted induction of acetogenesis into the rumen fermentation in vitro. Reprod. Nutr. Dev. 1996, 36, 233–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garsa, A.K.; Choudhury, P.K.; Puniya, A.K.; Dhewa, T.; Malik, R.K.; Tomar, S.K. Bovicins: The bacteriocins of streptococci and their potential in methane mitigation. Probiotics Antimicrob. Proteins 2019, 11, 1403–1413. [Google Scholar] [CrossRef]
- Goopy, J.P. Creating a low enteric methane emission ruminant: What is the evidence of success to the present and prospects for developing economies? Anim. Prod. Sci. 2019, 59, 1769–1776. [Google Scholar] [CrossRef] [Green Version]
- Hook, S.E.; Wright, A.D.G.; McBride, B.W. Methanogens: Methane producers of the rumen and mitigation strategies. Archaea 2010, 2010, 945785. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Mamuad, L.L.; Islam, M.; Lee, S.S. Reductive acetogens isolated from ruminants and their effect on in vitro methane mitigation and milk performance in Holstein cows. J. Anim. Sci. Technol. 2020, 62, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.; Morgavi, D.P.; Doreau, M. Methane mitigation in ruminants: From microbe to the farm scale. Animal 2010, 4, 351–365. [Google Scholar] [CrossRef] [Green Version]
- Wanapat, M.; Kongmun, P.; Poungchompu, O.; Cherdthong, A.; Khejornsart, P.; Pilajun, R.; Kaenpakdee, S. Effects of plants containing secondary compounds and plant oils on rumen fermentation and ecology. Trop. Anim. Health Prod. 2012, 44, 399–405. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Kreuzer, M.O.; O’Mara, F.P.; McAllister, T.A. Nutritional management for enteric methane abatement: A review. Aust. J. Exp. Agric. 2008, 48, 21–27. [Google Scholar] [CrossRef]
- García-González, R.; López, S.; Fernández, M.; Bodas, R.; González, J.S. Screening the activity of plants and spices for decreasing ruminal methane production in vitro. Anim. Feed Sci. Technol. 2008, 147, 36–52. [Google Scholar] [CrossRef]
- Eckard, R.J.; Grainger, C.; de Klein, C.A.M. Options for the abatement of methane and nitrous oxide from ruminant production: A review. Livest. Sci. 2011, 130, 47–56. [Google Scholar] [CrossRef]
- Patra, A.; Park, T.; Kim, M.; Yu, Z. Rumen methanogens and mitigation of methane emission by anti-methanogenic compounds and substances. J. Anim. Sci. Biotechnol. 2017, 8, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGinn, S.M.; Beauchemin, K.A.; Coates, T.; Colombatto, D. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast and furmaric acid. J. Anim. Sci. 2004, 82, 3346–3356. [Google Scholar] [CrossRef] [PubMed]
- Janssen, P.H.; Frenzel, P. Inhibition of methanogenesis by methyl fluoride: Studies of pure and defined mixed cultures of anaerobic bacteria and archaea. Appl. Environ. Microbiol. 1997, 63, 4552–4557. [Google Scholar] [CrossRef] [Green Version]
- Miller, T.L.; Wolin, M.J. Inhibition of growth of methane-producing bacteria of the ruminant forestomach by hydroxymethylglutaryl-SCoA reductase inhibitors. J. Dairy Sci. 2001, 84, 1445–1448. [Google Scholar] [CrossRef]
- Mitsumori, M.; Shinkai, T.; Takenaka, A.; Enishi, O.; Higuchi, K.; Kobayashi, Y.; Nonaka, I.; Asanuma, N.; Denman, S.E.; McSweeney, C.S. Responses in digestion, rumen fermentation and microbial populations to inhibition of methane formation by a halogenated methane analogue. Br. J. Nutr. 2012, 108, 482–491. [Google Scholar] [CrossRef]
- Sanchez, J.M.; Valle, L.; Rodriguez, F.; Morinnigo, M.A.; Borrego, J.J. Inhibition of methanogenesis by several heavy metals using pure cultures. Lett. Appl. Microbiol. 1996, 23, 439–444. [Google Scholar] [CrossRef]
- Fievez, V.F.; Dohme, M.; Daneels, K.R.; Demeyer, D. Fish oils as potent rumen methane inhibitors and associated effects on rumen fermentation in vitro and in vivo. Anim. Feed Sci. Technol. 2003, 104, 41–58. [Google Scholar] [CrossRef]
- Machmüller, A.; Kreuzer, M. Methane suppression by coconut oil and associated effects on nutrient and energy balance in sheep. Can. J. Anim. Sci. 1999, 79, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Machmüller, A. Medium-chain fatty acids and their potential to reduce methanogenesis in domestic ruminants. Agric. Ecosyst. Environ. 2006, 112, 107–114. [Google Scholar] [CrossRef]
- Lee, S.S.; Hsu, J.T.; Mantovani, H.C.; Russell, J.B. The effect of bovicin HC5, a bacteriocin from Streptococcus bovis HC5, on ruminal methane production in vitro. FEMS Microbiol. Lett. 2002, 217, 51–55. [Google Scholar] [CrossRef]
- Baker, S.K. Rumen methanogens and inhibition of methanogenesis. Aust. J. Agric. Res. 1999, 50, 1293–1298. [Google Scholar] [CrossRef]
- Anderson, R.C.; Carstens, G.E.; Miller, R.K.; Callaway, T.R.; Schultz, C.L.; Edrington, T.S.; Harvey, R.; Nisbet, D. Effect of oral nitroethane and 2-nitropropanol administration on methane-producing activity and volatile fatty acid production in the ovine rumen. Bioresour. Technol. 2006, 97, 2421–2426. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.D.G.; Kennedy, P.; O’neill, C.J.; Toovey, A.F.; Popovski, S.; Rea, S.M.; Pimm, C.L.; Klein, L. Reducing methane emissions in sheep by immunization against rumen methanogens. Vaccine 2004, 22, 3976–3985. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Huang, X.; Xue, B.; Peng, Q.; Wang, Z.; Yan, T.; Wang, L. Immunization against rumen methanogenesis by vaccination with a new recombinant protein. PLoS ONE 2015, 10, e0140086. [Google Scholar] [CrossRef] [Green Version]
- Newbold, C.J.; de la Fuente, G.; Belanche, A.; Ramos-Morales, E.; McEwan, N.R. The Role of Ciliate Protozoa in the Rumen. Front. Microbiol. 2015, 6, 1313. [Google Scholar] [CrossRef] [Green Version]
- LeVan, T.D.; Robinson, J.A.; John, R.; Greening, R.C.; Smolenski, W.J.; Leedle, J.A.Z.; Schaefer, D.M. Assessment of reductive acetogenesis with indigenous ruminal bacterium populations and Acetitomaculum ruminis. Appl. Environ. Microbiol. 1998, 64, 3429–3436. [Google Scholar] [CrossRef] [Green Version]
- Nollet, L.; Demeyer, D.; Verstraete, W. Effect of 2-bromoethanesulfonic acid and Peptostreptococcus productus ATCC35244 addition on stimulation of reductive acetogenesis in the ruminal ecosystem by selective inhibition of methanogenesis. Appl. Environ. Microbiol. 1997, 63, 194–200. [Google Scholar] [CrossRef] [Green Version]
- Nollet, L.; Velde, I.V.; Verstraete, W. Effect of the addition of Peptostreptococcus productus ATCC35244 on the gastro-intestinal microbiota and its activity, as simulated in an in vitro simulator of the human gastro-intestinal tract. Appl. Microbiol. Biotechnol. 1997, 48, 99–104. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; Wang, A.; Chen, J. Chemical inhibitors of methanogenesis and putative applications. Appl. Microbiol. Biotechnol. 2011, 89, 1333–1340. [Google Scholar] [CrossRef]
- Hungate, R.E. The Rumen and Its Microbes; Academic Press: New York, NY, USA, 1966; p. 533. [Google Scholar]
- Ungerfeld, E.M. Metabolic hydrogen flows in rumen fermentation: Principles and possibilities of interventions. Front. Microbiol. 2020, 11, 589. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.G.; Coleman, G.S. The rumen protozoa. In The Rumen Microbial Ecosystem, 2nd ed.; Hobson, P.N., Stewarteds, C.S., Eds.; Blackie Academic & Professional: New York, NY, USA, 1997; pp. 73–139. [Google Scholar]
- Cord-Ruwisch, R.; Seitz, H.J.; Conrad, R. The capacity of hydrogenotrophic anaerobic bacteria to compete for traces of hydrogen depends on the redox potential of the electron acceptor. Arch. Microbiol. 1988, 149, 350–357. [Google Scholar] [CrossRef]
- Immig, I. The rumen and hindgut as a source of ruminant methanogenesis. Environ. Monit. Assess. 1996, 42, 57–72. [Google Scholar] [CrossRef] [PubMed]
- McAllister, T.A.; Newbold, C.J. Redirecting rumen fermentation to reduce methanogenesis. Aust. J. Exp. Agric. 2008, 48, 7–13. [Google Scholar] [CrossRef]
- Ellis, J.L.; Dijkstra, J.; Kebreab, E.; Bannink, A.; Odongo, N.E.; Mcbride, B.W.; France, J. Aspects of rumen microbiology central to mechanistic modelling of methane production in cattle. J. Agric. Sci. 2008, 146, 213–233. [Google Scholar] [CrossRef] [Green Version]
- Hegarty, R.S.; Gerdes, R. Hydrogen production and transfer in the rumen. Rec. Adv. Anim. Nutr. 1999, 12, 37–44. Available online: http://livestocklibrary.com.au/handle/1234/19891 (accessed on 25 October 2022).
- Greening, C.; Geier, R.; Wang, C.; Woods, L.C.; Morales, S.E.; McDonald, M.J.; Rushton-Green, R.; Morgan, X.C.; Koike, S.; Leahy, S.C.; et al. Diverse hydrogen production and consumption pathways influence methane production in ruminants. ISME J. 2019, 13, 2617–2632. [Google Scholar] [CrossRef]
- Morgavi, D.P.; Forano, E.; Martin, C.; Newbold, C.J. Microbial ecosystem and methanogenesis in ruminants. Animal 2010, 4, 1024–1036. [Google Scholar] [CrossRef] [Green Version]
- Gagen, E.J.; Denman, S.E.; McSweeney, C.S. Acetogenesis as an alternative to methanogenesis in the rumen. In Livestock Production and Climate Change; Malik, P.K., Bhatta, R., Takahashi, J., Kohn, R.A., Prasad, C.S., Eds.; CABI: Wallingford, UK, 2015; pp. 292–303. [Google Scholar]
- Morvan, B.; Bonnemoy, F.; Fonty, G.; Gouet, P. Quantitative determination of H2-utilizing acetogenic and sulfate-reducing bacteria and methanogenic archaea from digestive tract of different mammals. Curr. Microbiol. 1996, 32, 129–133. [Google Scholar] [CrossRef]
- Kim, S.H.; Mamuad, L.L.; Kim, D.W.; Kim, S.K.; Lee, S.S. Fumarate reductase-producing enterococci reduce methane production in rumen fermentation in vitro. J. Microbiol. Biotechnol. 2016, 26, 558–566. [Google Scholar] [CrossRef]
- Mamuad, L.L.; Kim, S.H.; Lee, S.S.; Cho, K.K.; Jeon, C.O.; Lee, S.S. Characterization, metabolites and gas formation of fumarate reducing bacteria isolated from Korean native goat (Capra hircus coreanae). J. Microbiol. 2012, 50, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Wolin, M.J.; Miller, T.L.; Stewart, C.S. Microbe-microbe interactions. In The Rumen Microbial Ecosystem; Hobson, P.N., Stewart, C.S., Eds.; Chapman and Hall: London, UK, 1997; pp. 467–488. [Google Scholar]
- Joblin, K.N. Ruminal acetogens and their potential to lower ruminant methane emissions. Aust. J. Agric. Res. 1999, 50, 1307–1313. [Google Scholar] [CrossRef]
- Ljungdahl, L.G. The autotrophic pathway of acetate synthesis in acetogenic bacteria. Annu. Rev. Microbiol. 1986, 40, 415–450. [Google Scholar] [CrossRef] [PubMed]
- Morvan, B.; Doré, J.; Rieu-Lesme, F.; Foucat, L.; Fonty, G.; Gouet, P. Establishment of hydrogen-utilizing bacteria in the rumen of the newborn lamb. FEMS Microbiol. Lett. 1994, 117, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Ragsdale, S.W. Enzymology of the Wood Ljungdahl pathway of acetogenesis. Ann. N. Y. Acad. Sci. 2008, 1125, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Attwood, G.; McSweeney, C. Methanogen genomics to discover targets for methane mitigation technologies and options for alternative hydrogen utilization in the rumen. Aust. J. Exp. Agric. 2008, 48, 28–37. [Google Scholar] [CrossRef]
- Drake, H.L.; Küsel, K.; Matthies, C. Acetogenic prokaryotes. In The Prokaryotes: A Handbook on the Biology of Bacteria: Ecophysiology and Biochemistry; Dworkin, M., Stanley, F., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 354–420. [Google Scholar]
- Drake, H.L.; Gössner, S.A.; Daniel, S.L. Old acetogens, new light. Ann. N. Y. Acad. Sci. 2008, 1125, 100–128. [Google Scholar] [CrossRef]
- Klieve, A.V.; Ouwerkerk, D. Comparative greenhouse gas emissions from herbivores. In Proceedings of the VII International Symposium on the Nutrition of Herbivores; Meng, Q.X., Ed.; China Agricultural University Press: Beijing, China, 2007; pp. 17–21. [Google Scholar]
- Ungerfeld, E.M.; Kohn, R.A. The role of thermodynamics in the control of ruminal fermentation. In Ruminant Physiology: Digestion, Metabolism and Impact of Nutrition on Gene Expression, Immunology and Stress; Sejrsen, K., Hvelplund, T., Nielsen, M.O., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2006; pp. 55–85. [Google Scholar]
- Breznak, J.A.; Switzer, J.M. Acetate synthesis from H2 plus CO2 by termite gut microbes. Appl. Environ. Microbiol. 1986, 52, 623–630. [Google Scholar] [CrossRef] [Green Version]
- Breznak, J.A.; Kane, M.D. Microbial H2/CO2 acetogenesis in animal guts: Nature and nutritional significance. FEMS Microbiol. Rev. 1990, 87, 309–314. [Google Scholar] [CrossRef]
- Van Zijderveld, S.M.; Gerrits, W.J.; Apajalahti, J.A.; Newbold, J.R.; Dijkstra, J.; Leng, R.A.; Perdok, H. Nitrate and sulfate: Effective alternative hydrogen sinks for mitigation of ruminal methane production in sheep. J. Dairy Sci. 2010, 93, 5856–5866. [Google Scholar] [CrossRef] [Green Version]
- Silivong, P.; Preston, T.R.; Leng, R.A. Effect of sulphur and calcium nitrate on methane production by goats fed a basal diet of molasses supplemented with mimosa (Mimosa pigra) foliage. Livest. Res. Rural Dev. 2011, 23, 3. Available online: http://www.lrrd.org/lrrd23/3/sili23058.htm (accessed on 25 October 2022).
- Wu, H.; Meng, Q.; Yu, Z. Effect of pH buffering capacity and sources of dietary sulfur on rumen fermentation, sulfide production, methane production, sulfate reducing bacteria, and total Archaea in in vitro rumen cultures. Bioresour. Technol. 2015, 186, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Binversie, E.Y.; Ruiz-Moreno, M.; Carpenter, A.; Heins, B.; Crawford, G.; DiCostanzo, A.; Stern, M. Effects of dietary roughage and sulfur in diets containing corn dried distillers’ grains with solubles on hydrogen sulfide production and fermentation by rumen microbes in vitro. J. Anim. Sci. 2016, 94, 3883–3893. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.M.; Ma, J.; Wang, Z.; Hu, R.; Wang, X.; Peng, Q.; Amevor, F.K.; Goswami, N. Production of hydrogen sulfide by fermentation in rumen and its impact on health and production of animals. Processes 2020, 8, 1169. [Google Scholar] [CrossRef]
- Drewnoski, M.E.; Richter, E.L.; Hansen, S.L. Dietary sulfur concentration affects rumen hydrogen sulfide concentrations in feedlot steers during transition and finishing. J. Anim. Sci. 2012, 90, 4478–4486. [Google Scholar] [CrossRef]
- Gagen, E.J.; Mosoni, P.; Denman, S.E.; Al Jassim, R.; McSweeney, C.S.; Forano, E. Methanogen colonisation does not significantly alter acetogen diversity in lambs isolated 17 h after birth and raised aseptically. Microb. Ecol. 2012, 64, 628–640. [Google Scholar] [CrossRef]
- Fonty, G.; Joblin, K.; Chavarot, M.; Roux, R.; Naylor, G.; Michallon, F. Establishment and development of ruminal hydrogenotrophs in methanogen-free lambs. Appl. Environ. Microbiol. 2007, 73, 6391–6403. [Google Scholar] [CrossRef] [Green Version]
- Sakthivel, P.C.; Kamra, D.N.; Agarwal, N.; Chaudhary, L.C. Effect of sodium nitrate and nitrate reducing bacteria on in vitro methane production and fermentation with buffalo rumen liquor. Asian-Australas. J. Anim. Sci. 2012, 25, 812–817. [Google Scholar] [CrossRef] [Green Version]
- Simon, J. Enzymology and bioenergetics of respiratory nitrite ammonification. FEMS Microbiol. Rev. 2002, 26, 285–309. [Google Scholar] [CrossRef]
- Leng, R.A. The potential of feeding nitrate to reduce enteric methane production in ruminants. In Report: The Department of Climate Change; Commonwealth Government of Australia: Canberra, Australia, 2008. [Google Scholar]
- Takahashi, J. Some prophylactic options to mitigate methane emission from animal agriculture in Japan. Asian-Australas. J. Anim. Sci. 2011, 24, 285–294. [Google Scholar] [CrossRef]
- Van Zijderveld, S.M.; Fonken, B.; Dijkstra, J.; Gerrits, W.J.; Perdok, H.B.; Fokkink, W.; Newbold, J. Effects of a combination of feed additives on methane production, diet digestibility, and animal performance in lactating dairy cows. J. Dairy Sci. 2011, 94, 1445–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Rooke, J.A.; Cabeza, I.; Wallace, R.J. Nitrate and inhibition of ruminal methanogenesis: Microbial ecology, obstacles, and opportunities for lowering methane emissions from ruminant livestock. Front. Microbiol. 2016, 7, 132. [Google Scholar] [CrossRef]
- Iwamoto, M.; Asanuma, N.; Hino, T. Ability of Selenomonas ruminantium, Veillonella parvula and Wolinella succinogenes to reduce nitrate and nitrite with special reference to the suppression of ruminal methanogenesis. Anaerobe 2002, 8, 209–215. [Google Scholar] [CrossRef]
- Zhao, L.; Meng, Q.; Ren, L.; Liu, W.; Zhang, X.; Huo, Y.; Zhou, Z. Effects of nitrate addition on rumen fermentation, bacterial biodiversity and abundance. Asian-Australas. J. Anim. Sci. 2015, 28, 1433–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asanuma, N.; Yokoyama, S.; Hino, T. Effects of nitrate addition to a diet on fermentation and microbial populations in the rumen of goats, with special reference to Selenomonas ruminantium having the ability to reduce nitrate and nitrite. Anim. Sci. J. 2015, 86, 378–384. [Google Scholar] [CrossRef]
- Sar, C.; Santoso, B.; Mwenya, B.; Gamo, Y.; Kobayashi, T.; Morikawa, R.; Kimura, K.; Mizukoshi, H.; Takahashi, J. Manipulation of rumen methanogenesis by the combination of nitrate with β 1-4 galacto-oligosaccharides or nisin in sheep. Anim. Feed Sci. Technol. 2004, 115, 129–142. [Google Scholar] [CrossRef]
- Hulshof, R.B.; Berndt, A.; Gerrits, W.J.; Dijkstra, J.; van Zijderveld, S.M.; Newbold, J.R.; Perdok, H.B. Dietary nitrate supplementation reduces methane emission in beef cattle fed sugarcane-based diets. J. Anim. Sci. 2012, 90, 2317–2323. [Google Scholar] [CrossRef]
- Van Zijderveld, S.M.; Gerrits, W.J.; Dijkstra, J.; Newbold, J.R.; Hulshof, R.B.; Perdok, H.B. Persistency of methane mitigation by dietary nitrate supplementation in dairy cows. J. Dairy Sci. 2011, 94, 4028–4038. [Google Scholar] [CrossRef] [Green Version]
- Nolan, J.V.; Hegarty, R.S.; Hegarty, J.; Godwin, I.R.; Woodgate, R. Effects of dietary nitrate on fermentation, methane production and digesta kinetics in sheep. Anim. Prod. Sci. 2010, 50, 801–806. [Google Scholar] [CrossRef]
- Zhou, Z.; Yu, Z.; Meng, Q. Effects of nitrate on methane production, fermentation, and microbial populations in in vitro ruminal cultures. Bioresour. Technol. 2012, 103, 173–179. [Google Scholar] [CrossRef]
- Janssen, P.H. Influence of hydrogen on rumen methane formation and fermentation balances through microbial growth kinetics and fermentation thermodynamics. Anim. Feed Sci. Technol. 2010, 160, 1–22. [Google Scholar] [CrossRef]
- Chen, J.; Harstad, O.M.; McAllister, T.; Dörsch, P.; Holo, H. Propionic acid bacteria enhance ruminal feed degradation and reduce methane production in vitro. Acta Agric. Scand. Sect. A-Anim. Sci. 2020, 69, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Newbold, C.J.; López, S.; Nelson, N.; Ouda, J.O.; Wallace, R.J.; Moss, A.R. Propionate precursors and other metabolic intermediates as possible alternative electron acceptors to methanogenesis in ruminal fermentation in vitro. Br. J. Nutr. 2005, 94, 27–35. [Google Scholar] [CrossRef] [PubMed]
- López, S.; Valde’s, C.; Newbold, C.J.; Wallace, R.J. Influence of sodium fumarate addition on rumen fermentation in vitro. Br. J. Nutr. 1999, 81, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Bayaru, E.; Kanda, S.; Kamada, T.; Itabashi, H.; Andoh, S.; Nishida, T.; Ishida, M.; Itoh, T.; Nagara, K.; Isobe, Y. Effect of fumaric acid on methane production, rumen fermentation and digestibility of cattle fed roughage alone. Anim. Sci. J. 2001, 72, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Mao, S.Y.; Zhang, G.; Zhu, W.Y. Effect of disodium fumarate on ruminal metabolism and rumen bacterial communities as revealed by denaturing gradient gel electrophoresis analysis of 16S ribosomal DNA. Anim. Feed Sci. Technol. 2008, 140, 293–306. [Google Scholar] [CrossRef]
- Lin, B.; Lu, Y.; Salem, A.Z.M.; Wang, J.H.; Liang, Q.; Liu, J.X. Effects of essential oil combinations on sheep ruminal fermentation and digestibility of a diet with fumarate included. Anim. Feed Sci. Technol. 2013, 184, 24–32. [Google Scholar] [CrossRef]
- Foley, P.A.; Kenny, D.A.; Lovett, D.K.; Callan, J.J.; Boland, T.M.; O’Mara, F.P. Effect of DL-malic acid supplementation on feed intake, methane emissions, and performance of lactating dairy cows at pasture. J. Dairy Sci. 2009, 92, 3258–3264. [Google Scholar] [CrossRef] [Green Version]
- Foley, P.; Kenny, D.; Callan, J.; Boland, T.; O’mara, F. Effect of DL-malic acid supplementation on feed intake, methane emission, and rumen fermentation in beef cattle. J. Anim. Sci. 2009, 87, 1048–1057. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.W.; McSweeney, C.S.; Wang, J.K.; Liu, J.X. Effects of disodium fumarate on ruminal fermentation and microbial communities in sheep fed on high-forage diets. Animal 2012, 6, 815–823. [Google Scholar] [CrossRef]
- Bannink, A.; Kogut, J.; Dijkstra, J.; France, J.; Kebreab, E.; Van Vuuren, A.M.; Tamminga, S. Estimation of the stoichiometry of volatile fatty acid production in the rumen of lactating cows. J. Theor. Biol. 2006, 238, 36–51. [Google Scholar] [CrossRef] [PubMed]
- Ungerfeld, E.M. A theoretical comparison between two ruminal electron sinks. Front. Microbiol. 2013, 4, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackie, R.I.; Bryant, M.P. Acetogenesis and the rumen: Syntrophic relationships. In Acetogenesis; Drake, H.L., Ed.; Chapman and Hall: New York, NY, USA, 1994; pp. 331–364. [Google Scholar]
- Pinder, R.S.; Patterson, J.A. Growth of acetogenic bacteria in response to varying pH, acetate or carbohydrate concentration. Agric. Food Anal. Bacteriol. 2012, 3, 6–16. [Google Scholar]
- López, S.; McIntosh, F.M.; Wallace, R.J.; Newbold, C.J. Effect of adding acetogenic bacteria on methane production by mixed rumen microorganisms. Anim. Feed Sci. Technol. 1999, 78, 1–9. [Google Scholar] [CrossRef]
- Mitsumori, M.; Matsui, H.; Tajima, K.; Shinkai, T.; Takenaka, A.; Denman, S.E.; McSweeney, C.S. Effect of bromochloromethane and fumarate on phylogenetic diversity of the formyltetrahydrofolate synthetase gene in bovine rumen. Anim. Sci. J. 2014, 85, 25–31. [Google Scholar] [CrossRef]
- Faichney, G.J.; Graham, N.M.; Walker, D.M. Rumen characteristics, methane emission, and digestion in weaned lambs reared in isolation. Aust. J. Agric. Res. 1999, 50, 1083–1090. [Google Scholar] [CrossRef]
- Chaucheyras, F.; Fonty, G.; Bertin, G.; Gouet, P. In vitro H2 utilization by a ruminal acetogenic bacterium cultivated alone or in association with an archaea methanogen is stimulated by a probiotic strain of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1995, 61, 3466–3467. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Guan, L.; Liu, J.; Wang, J. Rumen fermentation and acetogen population changes in response to an exogenous acetogen TWA4 strain and Saccharomyces cerevisiae fermentation product. J. Zhejiang Univ. Sci. B 2015, 16, 709–719. [Google Scholar] [CrossRef] [Green Version]
- Ouwerkerk, D.; Maguire, A.; McMillen, L.; Klieve, A.V. Hydrogen utilising bacteria from the forestomach of eastern grey (Macropus giganteus) and red (Macropus rufus) kangaroos. Anim. Prod. Sci. 2009, 49, 1043–1051. [Google Scholar] [CrossRef]
- Gagen, E.J.; Wang, J.; Padmanabha, J.; Liu, J.; de Carvalho, I.P.C.; Liu, J.; Webb, R.; Al Jassim, R.; Morrison, M.; E Denman, S.; et al. Investigation of a new acetogen isolated from an enrichment of the tammar wallaby forestomach. BMC Microbiol. 2014, 14, 314. [Google Scholar] [CrossRef] [Green Version]
- Breznak, J.A. Acetogenesis from carbon dioxide in termite guts. In Acetogenesis; Drake, H.L., Ed.; Chapman and Hall: New York, NY, USA, 1994; pp. 303–330. [Google Scholar]
- Klieve, A.V.; Joblin, K.N. Comparison in hydrogen utilization of ruminal and marsupial reductive acetogens. In Proceedings of the PGGRC 5-Year Science Progress Report, 3rd International Greenhouse Gases and Animal Agriculture Conference, Christchurch, New Zealand, 26–29 November 2007; pp. 34–35. [Google Scholar]
- Short, S.B.; Edwards, W.C. Sulfur (hydrogen-sulfide) toxicosis in cattle. Vet. Hum. Toxicol. 1989, 31, 451–453. [Google Scholar] [PubMed]
- Odom, J.M.; Singleton, R., Jr. The Sulfate-Reducing Bacteria: Contemporary Perspectives; Springer-Verlag Inc.: New York, NY, USA, 1993; p. 289. [Google Scholar] [CrossRef]
- Thomas, W.E.; Loosli, J.K.; Williams, H.H.; Maynard, L.A. The utilization of inorganic sulfates and urea nitrogen by lambs. J. Nutr. 1951, 43, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Huisingh, J.; McNeill, J.J.; Matrone, G. Sulphate reduction by a Desulfovibrio species isolated from sheep rumen. Appl. Microbiol. 1974, 28, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Cummings, B.A.; Caldwell, D.R.; Gould, D.H.; Hamar, D.W. Identity and interactions of rumen microbes associated with dietary sulfate-induced polioencephalomalacia. Am. J. Vet. Res. 1995, 56, 1384–1389. [Google Scholar] [PubMed]
- Kushkevych, I.; Kotrsová, V.; Dordević, D.; Buňková, L.; Vítězová, M.; Amedei, A. Hydrogen sulfide effects on the survival of lactobacilli with emphasis on the development of inflammatory bowel diseases. Biomolecules 2019, 9, 752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, S.; Mu, C.; Zhu, W. Diversity and community pattern of sulfate-reducing bacteria in piglet gut. J. Anim. Sci. Biotechnol. 2019, 10, 40. [Google Scholar] [CrossRef]
- McCann, J.C.; Segers, J.R.; Derakhshani, H.; Felix, T.L.; Khafipour, E.; Shike, D.W. Increasing corn distillers solubles alters the liquid fraction of the ruminal microbiome. J. Anim. Sci. 2017, 95, 3540–3551. [Google Scholar] [CrossRef]
- Howard, B.H.; Hungate, R.E. Desulfovibrio of the sheep rumen. Appl. Environ. Microbiol. 1976, 32, 598–602. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.S.; Deb, S.M.; Singh, D. Isolation and characterization of novel sulphate-reducing Fusobacterium sp. and their effects on in vitro methane emission and digestion of wheat straw by rumen fluid from Indian riverine buffaloes. Anim. Feed Sci. Technol. 2011, 166, 132–140. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Sheep, 6th ed.; National Academy Press: Washington, DC, USA, 1985. [Google Scholar]
- NRC. Nutrient Requirements of Beef Cattle, 7th ed.; National Academy Press: Washington, DC, USA, 2000. [Google Scholar]
- Whanger, P.D.; Matrone, G. Effect of dietary sulfur upon the production and absorption of lactate in sheep. Biochim. Biophys. Acta 1996, 124, 273–279. [Google Scholar] [CrossRef]
- Rumsey, T.S. Effects of dietary sulfur addition and Synovex-S ear implants on feedlot steers fed an all-concentrate finishing diet. J. Anim. Sci. 1978, 46, 463–477. [Google Scholar] [CrossRef]
- Whanger, P.D.; Matrone, G. Effect of dietary sulfur upon the fatty acid production in the rumen. Biochim. Biophys. Acta 1965, 98, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Hegarty, R.S.; Nolan, J.V.; Leng, R.A. The effects of protozoa and of supplementation with nitrogen and sulfur on digestion and microbial metabolism in the rumen of sheep. Aust. J. Agric. Res. 1994, 45, 1215–1227. [Google Scholar] [CrossRef]
- Patterson, J.A.; Kung, L. Metabolism of DL-methionine and methionine analogs by rumen micro-organisms. J. Dairy Sci. 1988, 71, 3292–3301. [Google Scholar] [CrossRef] [PubMed]
- Inthapanya, S.; Preston, T.R.; Khang, D.N.; Leng, R.A. Effect of potassium nitrate and urea as fermentable nitrogen sources on growth performance and methane emissions in local “Yellow” cattle fed lime (Ca(OH)2) treated rice straw supplemented with fresh cassava foliage. Livest. Res. Rural Dev. 2012, 24, 27. Available online: http://www.lrrd.org/lrrd24/2/sang24027.htm (accessed on 25 October 2022).
- Sophal, C.; Khang, D.N.; Preston, T.R.; Leng, R.A. Nitrate replacing urea as a fermentable N source decreases enteric methane production and increases the efficiency of feed utilization in Yellow cattle. Livest. Res. Rural Dev. 2013, 25, 113. Available online: http://www.lrrd.org/lrrd25/7/soph25113.htm (accessed on 25 October 2022).
- Ascensão, A.M.D. Effects of Nitrate and Additional Effect of Probiotic on Methane Emissions and Dry Matter Intake in Nellore Bulls. Master’s Thesis, Universidade de Trás-os-Montes e Alto Douro Departamento de Zootecnia, Vila Real, Portugal, 2010. [Google Scholar]
- Wood, T.A.; Wallace, R.J.; Rowe, A.; Price, J.; Yáñez-Ruiz, D.R.; Murray, P.; Newbold, C.J. Encapsulated fumaric acid as a feed ingredient to decrease ruminal methane emissions. Anim. Feed Sci. Technol. 2009, 152, 62–71. [Google Scholar] [CrossRef]
- Demeyer, D.I.; Henderickx, H.K. Competitive inhibition of in vitro methane production by mixed rumen bacteria. Arch. Internat. Physiol. 1967, 75, 157–159. [Google Scholar]
- Kolver, E.S.; Aspin, P.W.; Jarvis, G.N.; Elborough, K.M.; Roche, J.R. Fumarate reduces methane production from pasture fermented in continuous culture. Proc. N. Z. Soc. Anim. 2004, 64, 155–159. [Google Scholar]
- Newbold, C.J.; Ouda, J.O.; Lopez, S.; Nelson, N.; Omed, H.; Wallace, R.J.; Moss, A.R. Propionate precursors as possible alternative electron acceptors to methane in ruminal fermentation. In Greenhouse Gases and Animal Agriculture; Takahashi, J., Youngeds, B.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2002; pp. 151–154. [Google Scholar]
- Iwamoto, M.; Asanuma, N.; Hino, T. Effects of nitrate combined with fumarate on methanogenesis, fermentation, and cellulose digestion by rumen microbes in vitro. Anim. Sci. J. 1999, 70, 471–478. [Google Scholar] [CrossRef] [Green Version]
- Asanuma, N.; Iwamoto, M.; Hino, T. Effect of the addition of fumarate on methane production by ruminal microorganisms in vitro. J. Dairy Sci. 1999, 82, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.; Sarwar, M.; Mehr-un-Nisa; Hayat, Z.; Younas, M. Effect of supplementary sodium nitrate and sulphur on methane production and growth rates in sheep and goats fed forage-based diet low in true protein. J. Anim. Plant Sci. 2016, 26, 69–78. [Google Scholar]
- Sarturi, J.O.; Erickson, G.E.; Klopfenstein, T.J.; Rolfe, K.M.; Buckner, C.D.; Luebbe, M.K. Impact of source of sulfur on ruminal hydrogen sulfide and logic for the ruminal available sulfur for reduction concept. J. Anim. Sci. 2013, 91, 3352–3359. [Google Scholar] [CrossRef]
- Bray, A.C. Sulphur metabolism in sheep. II. Absorption of inorganic sulphate and inorganic sulphide from sheeps rumen. Aust. J. Agric. Res. 1969, 20, 739–748. [Google Scholar] [CrossRef]
- Richter, E.L.; Drewnoski, M.E.; Hansen, S.L. Effect of increased dietary sulfur on beef steer mineral status, performance, and meat fatty acid composition. J. Anim. Sci. 2012, 90, 3945–3953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drewnoski, M.E.; Doane, P.; Hansen, S.L. Ferric citrate decreases ruminal hydrogen sulphide concentrations in feedlot cattle fed diets high in sulphate. Br. J. Nutr. 2014, 111, 261–269. [Google Scholar] [CrossRef] [Green Version]
- DiLorenzo, N.; Dahlen, C.R.; Diez-Gonzalez, F.; Lamb, G.C.; Larson, J.E.; DiCostanza, A. Effects of feeding polyclonal antibody preparations on rumen fermentation patterns, performance, and carcass characteristics of feedlot steers. J. Anim. Sci. 2008, 86, 3023–3032. [Google Scholar] [CrossRef] [PubMed]
- Latham, E.A.; Anderson, R.C.; Pinchak, W.E.; Nisbet, D.J. Insights on Alterations to the rumen ecosystem by nitrate and nitrocompounds. Front. Microbiol. 2016, 7, 228. [Google Scholar] [CrossRef]
- Payne, W.J. Reduction of nitrogenous oxides by microorganisms. Bacteriol. Rev. 1973, 37, 409–452. [Google Scholar] [CrossRef]
- Baar, C.; Eppinger, M.; Raddatz, G.; Simon, J.; Lanz, C.; Klimmek, O.; Nandakumar, R.; Gross, R.; Rosinus, A.; Keller, H.; et al. Complete genome sequence and analysis of Wolinella succinogenes. Proc. Natl. Acad. Sci. USA 2003, 100, 11690–11695. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.S.; Schaefer, D.M.; Guo, X.X.; Ren, L.P.; Meng, Q.X. Use of nitrate-nitrogen as a sole dietary nitrogen source to inhibit ruminal methanogenesis and to improve microbial nitrogen synthesis in vitro. Asian-Australas. J. Anim. Sci. 2009, 22, 542–549. [Google Scholar] [CrossRef]
- Brunet, R.C.; Garcia-Gil, L.J. Sulfide-induced dissimilatory nitrate reduction to ammonia in anaerobic freshwater sediments. FEMS Microbiol. Ecol. 1996, 21, 131–138. [Google Scholar] [CrossRef]
- Leng, R.A.; Nolan, J.V. Nitrogen metabolism in the rumen. J. Dairy Sci. 1984, 67, 1072–1089. [Google Scholar] [CrossRef] [PubMed]
- Cockburn, A.; Brambilla, G.; Fernandez, M.-L.; Arcella, D.; Bordajandi, L.R.; Cottrill, B.; van Peteghem, C.; Dorne, J.-L. Nitrite in feed: From animal health to human health. Toxicol. Appl. Pharmacol. 2013, 270, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Alaboudi, A.; Jones, G. Effect of acclimation to high nitrate intakes on some rumen fermentation parameters in sheep. Can. J. Anim. Sci. 1985, 65, 841–849. [Google Scholar] [CrossRef]
- Hao, T.P.; Quang, D.H.; Preston, T.R.; Leng, R.A. Nitrate as a fermentable nitrogen supplement for goats fed forage based diets low in true protein. Livest. Res. Rural Dev. 2009, 21. Available online: https://www.lrrd.org/lrrd21/1/trin21010.htm (accessed on 25 October 2022).
- Jones, G.A. Dissimilatory metabolism of nitrate by the rumen microbiota. Can. J. Microbiol. 1972, 18, 1783–1787. [Google Scholar] [CrossRef] [PubMed]
- Sar, C.; Mwenya, B.; Pen, B.; Takaura, K.; Morikawa, R.; Tsujimoto, A.; Kuwaki, K.; Isogai, N.; Shinzato, I.; Asakura, Y.; et al. Effect of ruminal administration of Escherichia coli wild type or a genetically modified strain with enhanced high nitrite reductase activity on methane emission and nitrate toxicity in nitrate-infused sheep. Br. J. Nutr. 2005, 94, 691–697. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.S.; Bryant, M.P. The rumen bacteria. In The Rumen Microbial Ecosystem; Hobson, P.N., Stewart, C.S., Eds.; Blackie Academic and Professional: London, UK, 1988; pp. 21–75. [Google Scholar]
- Sar, C.; Mwenya, B.; Santoso, B.; Takaura, K.; Morikawa, R.; Isogai, N.; Asakura, Y.; Toride, Y.; Takahashi, J. Effect of Escherichia coli wild type or its derivative with high nitrite reductase activity on in vitro ruminal methanogenesis and nitrate/nitrite reduction. J. Anim. Sci. 2005, 83, 644–652. [Google Scholar] [CrossRef]
- Sar, C.; Mwenya, B.; Santoso, B.; Takaura, K.; Morikawa, R.; Isogai, N.; Asakura, Y.; Toride, Y.; Takahashi, J. Effect of Escherichia coli W3110 on ruminal methanogenesis and nitrate/ nitrite reduction in vitro. Anim. Feed Sci. Technol. 2005, 118, 295–306. [Google Scholar] [CrossRef]
- Mwenya, B.; Zhou, X.; Santoso, B.; Sar, C.; Gamo, Y.; Kobayashi, T.; Takahashi, J. Effects of probiotic-vitacogen and β 1-4 galacto-oligosaccharides supplementation on methanogenesis and energy and nitrogen utilization in dairy cows. Asian-Australas. J. Anim. Sci. 2004, 17, 349–354. [Google Scholar] [CrossRef]
- Yoshida, J.; Nakamura, Y.; Nakamura, R. Effects of protozoal fraction and lactate on nitrate metabolism of microorganisms in sheep rumen. Jpn. J. Zootech. Sci. 1982, 53, 677–685. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.; Schaefer, D.M.; Guo, W.S.; Ren, L.P.; Meng, Q.X. Comparisons of in vitro nitrate reduction, methanogenesis, and fermentation acid profile among rumen bacterial, protozoal and fungal fractions. Asian-Australas. J. Anim. Sci. 2011, 24, 471–478. [Google Scholar] [CrossRef]
- Iwamoto, M.; Asanuma, N.; Hino, T. Effects of energy substrates on nitrate reduction and nitrate reductase activity in a ruminal bacterium, Selenomonas ruminantium. Anaerobe 2001, 7, 315–321. [Google Scholar] [CrossRef]
- Sophea, I.v.; Preston, T.R. Effect of different levels of supplementary potassium nitrate replacing urea on growth rates and methane production in goats fed rice straw, mimosa foliage and water spinach. Livest. Res. Rural Dev. 2011, 23, 71. Available online: http://www.lrrd.org/lrrd23/4/soph23071.htm (accessed on 25 October 2022).
- Li, L.; Davis, J.; Nolan, J.; Hegarty, R. An initial investigation on rumen fermentation pattern and methane emission of sheep offered diets containing urea or nitrate as the nitrogen source. Anim. Prod. Sci. 2012, 52, 653–658. [Google Scholar] [CrossRef]
- Zhou, Z.; Meng, Q.; Yu, Z. Effects of methanogenic inhibitors on methane production and abundances of methanogens and cellulolytic bacteria in in vitro ruminal cultures. Appl. Environ. Microbiol. 2011, 77, 2634–2639. [Google Scholar] [CrossRef] [Green Version]
- Ungerfeld, E.M. Shifts in metabolic hydrogen sinks in the methanogenesis-inhibited ruminal fermentation: A meta-analysis. Front. Microbiol. 2015, 6, 37. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Nan, X.; Chu, K.; Tong, J.; Yang, L.; Zheng, S.; Zhao, G.; Jiang, L.; Xiong, B. Shifts of hydrogen metabolism from methanogenesis to propionate production in response to replacement of forage fiber with non-forage fiber sources in diets in vitro. Front. Microbiol. 2018, 9, 2764. [Google Scholar] [CrossRef] [Green Version]
- Castillo, C.; Benedito, J.L.; Mendez, J.; Pereira, V.; Lopez-Alonso, M.; Miranda, M.; Hernández, J. Organic acids as a substitute for monensin in diets for beef cattle. Anim. Feed Sci. Technol. 2004, 115, 101–116. [Google Scholar] [CrossRef]
- Baldwin, R.L.; Allison, J.M. Rumen metabolism. J. Anim. Sci. 1983, 57, 461–477. [Google Scholar] [PubMed]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.A. Manipulation of ruminal fermentation with organic acids: A review. J. Anim. Sci. 1998, 76, 3123–3132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisbet, D.J.; Martin, S.A. Effects of fumarate, L-malate, and an Aspergillus oryzae fermentation extract on D-lactate utilization by the ruminal bacterium Selenomonas ruminantium. Curr. Microbiol. 1993, 26, 133–136. [Google Scholar] [CrossRef]
- García-Martínez, R.; Ranilla, M.J.; Tejido, M.L.; Carro, M.D. Effects of disodium fumarate on in vitro rumen microbial growth, methane production and fermentation of diets differing in their forage: Concentrate ratio. Br. J. Nutr. 2005, 94, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Ungerfeld, E.M.; Kohn, R.A.; Wallace, R.J.; Newbold, C.J. A meta-analysis of fumarate effects on methane production in ruminal batch cultures. J. Anim. Sci. 2007, 85, 2556–2563. [Google Scholar] [CrossRef]
- Castro-Montoya, J.; De Campeneere, S.; Van Ranst, G.; Fievez, V. Interactions between methane mitigation additives and basal substrates on in vitro methane and VFA production. Anim. Feed Sci. Technol. 2012, 176, 47–60. [Google Scholar] [CrossRef]
- Wallace, R.J.; Wood, T.A.; Rowe, A.; Price, J.; Yanez, D.R.; Williams, S.P.; Newbold, C. Encapsulated fumaric acid as a means of decreasing ruminal methane emissions. Int. Congr. Ser. 2006, 1293, 148–151. [Google Scholar] [CrossRef]
- Itabashi, H.; Bayaru, E.; Kanda, S.; Nishida, T.; Ando, S.; Ishida, M.; Itoh, T.; Isobe, Y.; Nagara, K.; Takei, K. Effect of Salinomycin (SL) or SL plus Fumaric Acid on rumen fermentation and methane production in cattle. Asian-Australas. J. Anim. Sci. 2000, 13, 287. [Google Scholar]
- Beauchemin, K.A.; McGinn, S.M. Methane emissions from beef cattle: Effects of fumaric acid, essential oil, and canola oil. J. Anim. Sci. 2006, 84, 1489–1496. [Google Scholar] [CrossRef]
- Martin, S.A.; Streeter, M.N. Effect of malate on in vitro mixed ruminal microorganism fermentation. J. Anim. Sci. 1995, 73, 2141–2145. [Google Scholar] [CrossRef]
- Devant, M.; Bach, A.; García, J.A. Effect of malate supplementation to dairy cows on rumen fermentation and milk production in early lactation. J. Appl. Anim. Res. 2007, 31, 169–172. [Google Scholar] [CrossRef]
- Kung, L., Jr.; Huber, J.T.; Krummerey, J.D.; Allison, L.; Cook, R.M. Influence of adding malic acid to dairy cattle rations on milk production, rumen volatile acids, digestibility, and nitrogen utilization. J. Dairy Sci. 1982, 65, 1170–1174. [Google Scholar] [CrossRef]
- Sanson, D.W.; Stallcup, O.T. Growth response and serum constituents of Holstein bulls fed malic acid. Nutr. Rep. Int. 1984, 30, 1261–1267. [Google Scholar]
- Callaway, T.R.; Martin, S.A. Effects of organic acid and monensin treatment on in vitro mixed ruminal microorganism fermentation of cracked corn. J. Anim. Sci. 1996, 74, 1982–1989. [Google Scholar] [CrossRef] [Green Version]
- Carro, M.D.; López, S.; Valdés, C.; Ovejero, F.J. Effect of D, L-malate on mixed ruminal microorganism fermentation using the rumen simulation technique (RUSITEC). Anim. Feed Sci. Technol. 1999, 79, 279–288. [Google Scholar] [CrossRef]
- Carro, M.D.; Ranilla, M.J. Effect of the addition of malate on in vitro rumen fermentation of cereal grains. Br. J. Nutr. 2003, 89, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carro, M.D.; Ranilla, M.J. Influence of different concentrations of disodium fumarate on methane production and fermentation of concentrate feeds by rumen micro-organisms in vitro. Br. J. Nutr. 2003, 90, 617–623. [Google Scholar] [CrossRef] [Green Version]
- Asanuma, N.; Hino, T. Activity and properties of fumarate reductase in ruminal bacteria. J. Gen. Appl. Microbiol. 2000, 46, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Mamuad, L.; Kim, S.H.; Jeong, C.D.; Choi, Y.J.; Jeon, C.O.; Lee, S.S. Effect of fumarate reducing bacteria on in vitro rumen fermentation, methane mitigation and microbial diversity. J. Microbiol. 2014, 52, 120–128. [Google Scholar] [CrossRef]
- Astuti, W.D.; Wiryawan, K.G.; Wina, E.; Widyastuti, Y.; Suharti, S.; Ridwan, R. Effects of selected Lactobacillus plantarum as probiotic on in vitro ruminal fermentation and microbial population. Pak. J. Nutr. 2018, 17, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef] [PubMed]
- Kaewpila, C.; Gunun, P.; Kesorn, P.; Subepang, S.; Thip-uten, S.; Cai, Y.; Pholsen, S.; Cherdthong, A.; Khota, W. Improving ensiling characteristics by adding lactic acid bacteria modifies in vitro digestibility and methane production of forage-sorghum mixture silage. Sci. Rep. 2021, 11, 1968. [Google Scholar] [CrossRef] [PubMed]
- Varnava, K.G.; Ronimus, R.S.; Sarojini, V. A review on comparative mechanistic studies of antimicrobial peptides against archaea. Biotechnol. Bioeng. 2017, 114, 2457–2473. [Google Scholar] [CrossRef]
- Jeyanathan, J.; Martin, C.; Morgavi, D.P. Screening of bacterial direct-fed microbials for their antimethanogenic potential in vitro and assessment of their effect on ruminal fermentation and microbial profiles in sheep. J. Anim. Sci. 2016, 94, 739–750. [Google Scholar] [CrossRef]
- Callaway, T.R.; Carneiro De Melo, A.M.S.; Russell, J.B. The effect of nisin and monensin on ruminal fermentations in vitro. Curr. Microbiol. 1997, 35, 90–96. [Google Scholar] [CrossRef]
- Santoso, B.; Mwenya, B.; Sar, C.; Gamo, Y.; Kobayashi, T.; Morikawa, R.; Kimura, K.; Mizukoshi, H.; Takahashi, J. Effects of supplementing galacto-oligosaccharides, Yucca schidigera or nisin on ruminal methanogenesis, nitrogen and energy metabolism in sheep. Livest. Prod. Sci. 2004, 91, 209–217. [Google Scholar] [CrossRef]
- Renuka; Puniya, M.; Sharma, A.; Malik, R.; Upadhyay, R.C.; Puniya, A.K. Influence of pediocin and enterocin on in-vitro methane, gas production and digestibility. Int. J. Curr. Microbiol. Appl. Sci. 2013, 2, 132–142. [Google Scholar]
- Cao, Y.; Takahashi, T.; Horiguchi, K.; Yoshida, N. Effect of adding lactic acid bacteria and molasses on fermentation quality and in vitro ruminal digestion of total mixed ration silage prepared with whole crop rice. Grassl. Sci. 2010, 56, 19–25. [Google Scholar] [CrossRef]
- Cao, Y.; Cai, Y.; Takahashi, T.; Yoshida, N.; Tohno, M.; Uegaki, R.; Nonaka, K.; Terada, F. Effect of lactic acid bacteria inoculant and beet pulp addition on fermentation characteristics and in vitro ruminal digestion of vegetable residue silage. J. Dairy Sci. 2011, 94, 3902–3912. [Google Scholar] [CrossRef] [Green Version]
- Huyen, N.T.; Martinez, I.; Pellikaan, W. Using lactic acid bacteria as silage inoculants or direct-fed microbials to improve in vitro degradability and reduce methane emissions in dairy cows. Agronomy 2020, 10, 1482. [Google Scholar] [CrossRef]
- Guo, G.C.Q.; Shen, C.; Liu, Q.; Zhang, S.; Shao, T.; Wang, C.; Wang, Y.; Xu, Q.; Huo, W. The effect of lactic acid bacteria inoculums on in vitro rumen fermentation, methane production, ruminal cellulolytic bacteria populations and cellulase activities of corn stover silage. J. Integr. Agric. 2020, 19, 838–847. [Google Scholar] [CrossRef]





| Dietary Supplements | Source and Level | Model | CH4 Reduction (%) | References |
|---|---|---|---|---|
| Sulfur | Sulfate (2.6%) | Sheep | 16 | [70] |
| Sodium sulfate (0.8%) | Goat | 14.2 | [71] | |
| Nitrate | Pottasium nitrate (4%) | Sheep | 23 | [90] |
| Pottasium nitrate (5%) | Cattle | 43 | [132] | |
| Pottasium nitrate (6%) | Cattle | 27 | [133] | |
| Nitrate (22 g/kg DM) | Cattle | 32 | [88] | |
| Nitrate (2.6%) | Sheep | 32 | [70] | |
| Calcium ammonium nitrate (2.84%) | Cattle | 41 | [134] | |
| Sodium nitrate (1.3 g/kg BW) | Sheep | 50.4 | [87] | |
| Nitrate (21 g/kg DM) | Cattle | 16 | [89] | |
| Calcium nitrate (3.8%/DM) | Goat | 23.2 | [71] | |
| Fumarate | Fumaric acid (2% DM) | Cattle | 23 | [96] |
| Encapsulated fumarate (10%) | Sheep | 76 | [135] | |
| Sodium fumarate (400 μM) | In vitro | 17 | [94] | |
| Sodium fumarate (500 μM) | In vitro | 60 | [136] | |
| Fumarate (3.5 g/L) | In vitro | 38 | [137] | |
| Sodium fumarate (6.2 mM) | In vitro | 17 | [95] | |
| Fumaric acid (8% DM) | Sheep | 12 | [138] | |
| Sodium acrylate | In vitro | 8 | [94] | |
| Sodium fumarate | In vitro | 17 | [94] | |
| Fumarate (10 mM) | In vitro | 17 | [139] | |
| Fumarate (30 mM) | In vitro | 11 | [140] | |
| Combinations | Sulfur (2.6%) + Nitrate (2.6%) | Sheep | 47 | [70] |
| Sodium sulfate (0.8%) + Calcium nitrate (3.8%) | Goat | 34.9 | [71] | |
| Sodium nitrate (1.3 g/kg BW) + GOS | Sheep | 52.9 | [87] | |
| Sodium nitrate (1.3 g/kg BW) + Nisin (3 mg/kg BW) | Sheep | 56.3 | [87] | |
| Sodium nitrate (5%) + Sulfur (0.4%) | Sheep | 19.6 | [141] | |
| Sodium nitrate (5%) + Sulfur (0.4%) | Goat | 18.2 | [141] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choudhury, P.K.; Jena, R.; Tomar, S.K.; Puniya, A.K. Reducing Enteric Methanogenesis through Alternate Hydrogen Sinks in the Rumen. Methane 2022, 1, 320-341. https://doi.org/10.3390/methane1040024
Choudhury PK, Jena R, Tomar SK, Puniya AK. Reducing Enteric Methanogenesis through Alternate Hydrogen Sinks in the Rumen. Methane. 2022; 1(4):320-341. https://doi.org/10.3390/methane1040024
Chicago/Turabian StyleChoudhury, Prasanta Kumar, Rajashree Jena, Sudhir Kumar Tomar, and Anil Kumar Puniya. 2022. "Reducing Enteric Methanogenesis through Alternate Hydrogen Sinks in the Rumen" Methane 1, no. 4: 320-341. https://doi.org/10.3390/methane1040024
APA StyleChoudhury, P. K., Jena, R., Tomar, S. K., & Puniya, A. K. (2022). Reducing Enteric Methanogenesis through Alternate Hydrogen Sinks in the Rumen. Methane, 1(4), 320-341. https://doi.org/10.3390/methane1040024