
Species Abundance Distributions Patterns between Tiankeng Forests and Nearby Non-Tiankeng Forests in Southwest China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Survey and Sampling
2.3. Data Analysis
2.4. Models Selection
- (1)
- Fisher’s log-series model
- (2)
- Log-normal model
- (3)
- Broken stick model
- (4)
- Niche preemption model
- (5)
- Zero-sum polynomials model
- (6)
- Volkov model
2.5. Fitting Tests for Models
3. Results
3.1. Species Composition of Tiankeng Forests and Nearby Non-Tiankeng Forests
3.2. SADs Pattern in Tiankeng and Non-Tiankeng Forests
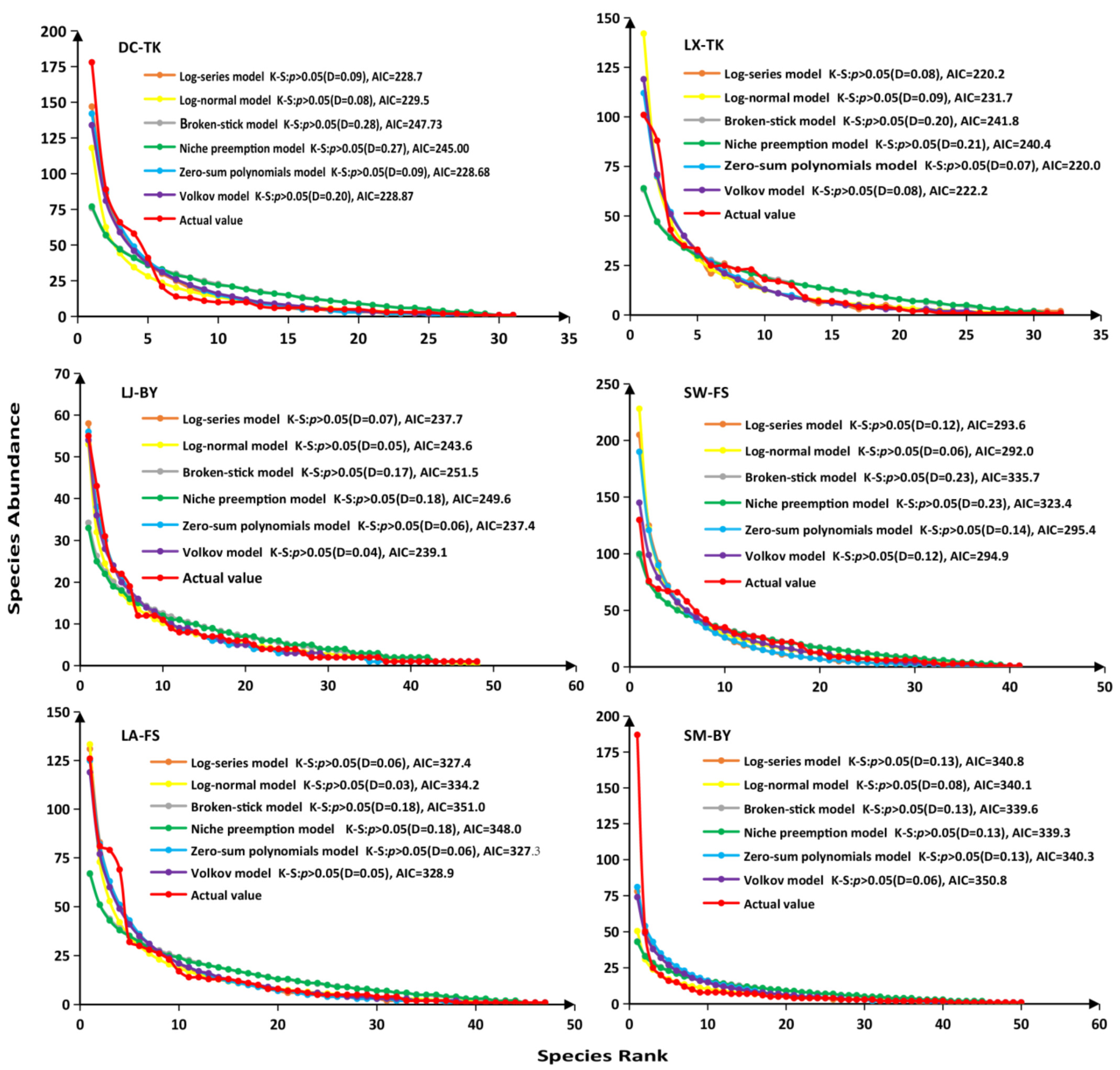
3.3. Goodness-of Fit of SADs Models in Tiankeng and Non-Tiankeng Forests
4. Discussion
4.1. Species Composition of Tiankeng Forest and Nearby Non-Tiankeng Forests
4.2. SADs Pattern in Tiankeng and Non-Tiankeng Forests
4.3. Goodness-of Fit of SADs Models in Tiankeng and Non-Tiankeng Forests
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dong, K.; Hao, G.; Yang, N.; Zhang, J.; Ding, X.; Ren, H.; Shen, J.; Wang, J.; Jiang, L.; Zhao, N.; et al. Community assembly mechanisms and succession processes significantly differ among treatments during the restoration of Stipa grandis–Leymus chinensis communities. Sci. Rep. 2019, 9, 16289. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, S.; Tysklind, N.; Derroire, G.; Heuertz, H.; Herault, B. Topography shapes the local coexistence of tree species within species complexes of Neotropical forests. Oecologia 2021, 196, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Germain, R.M.; Mayfield, M.; Gilbert, B. The ‘filtering’ metaphor revisited: Competition and environment jointly structure invasibility and coexistence. Biol. Lett. 2018, 14, 20180460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magura, T.; Lövei, G.L.; Tóthmérész, B. Conversion from environmental filtering to randomness as assembly rule of ground beetle assemblages along an urbanization gradient. Sci. Rep. 2018, 8, 16992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, D.; Ye, D.; Cadotte, M.W. Habitat loss-biodiversity relationships are influenced by assembly processes and the spatial configuration of area loss. For. Ecol. Manag. 2021, 496, 119452. [Google Scholar] [CrossRef]
- Matthews, T.J.; Whittaker, R.J. Neutral theory and the species abundance distribution: Recent developments and prospects for unifying niche and neutral perspectives. Ecol. Evol. 2014, 4, 2263–2277. [Google Scholar] [CrossRef] [PubMed]
- Kusumoto, B.; Kubota, Y.; Baselga, A.; Gómez, C.; Odríguez, O.C.; Shiono, T. Community dissimilarity of angiosperm trees reveals deep-time diversification across tropical and temperate forests. J. Veg. Sci. 2021, 32, e13017. [Google Scholar] [CrossRef]
- Ma, K.M. Advances of the Study on Species Abundance Pattern. Chin. J. Plant Ecol. 2003, 3, 412–426. Available online: https://www.plant-ecology.com/CN/10.17521/cjpe.2003.0060 (accessed on 15 September 2021).
- Hidasi, N.J.; Bini, L.M.; Siqueira, T.; Cianciaruso, M.V. Ecological similarity explains species abundance distribution of small mammal communities. Acta Oecol. 2020, 102, 103502. [Google Scholar] [CrossRef]
- Waldock, C.; Stuart-Smith, R.D.; Albouy, C.; Cheung, W.; Edgar, G.J.; Mouillot, D.; Tjiputra, J.; Pellissier, L. A quantitative review of abundance-based species distribution models. Ecography 2022, 1, e05694. [Google Scholar] [CrossRef]
- Thierry, E.H. Stochastic species abundance models involving special copulas. Phys. A. 2018, 490, 77–91. [Google Scholar] [CrossRef]
- Latimer, A.; Wu, S.; Silander, G. Building statistical models to analyze species distributions. Ecol. Appl. 2006, 16, 33–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takolander, A.; Hickler, T.; Meller, L.; Caneza, M. Comparing future shifts in tree species distributions across Europe projected by statistical and dynamic process-based models. Reg. Environ. Chang. 2019, 19, 251–266. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Tang, D. Estimating the niche preemption parameter of the geometric series. Acta Oecologica 2008, 33, 105–107. [Google Scholar] [CrossRef]
- Dallas, T.; Hastings, A. Habitat suitability estimated by niche models is largely unrelated to species abundance. Glob. Ecol. Biogeogr. 2018, 27, 1448–1456. [Google Scholar] [CrossRef]
- Barabás, G.; Andrea, R.; Rael, R.; Meszéna, G.; Ostling, A. Emergent neutrality or hidden niches. Oikos 2013, 122, 1565–1572. [Google Scholar] [CrossRef] [Green Version]
- Petr, D.; Karolina, T.; Tereza, K. Linking species abundance and overyielding from experimental communities with niche and fitness characteristics. J. Ecol. 2019, 107, 178–189. [Google Scholar] [CrossRef]
- Volkov, I.; Banavar, J.R.; Hubbell, S.P.; Maritan, A. Neutral theory and relative species abundance in ecology. Nature 2003, 424, 1035–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.Z.; Ellwood, M.; Ai, D.; Zhang, R.Y.; Wang, G. Species abundance distributions as a proxy for the niche-neutrality continuum. J. Plant Ecol. 2018, 11, 445–452. [Google Scholar] [CrossRef]
- Villa, P.M.; Martins, S.V.; Rodrigues, A.C.; Alice, C.R.; Nathália, V.H.; Michael, A.C.; Ali, A. Testing species abundance distribution models in tropical forest successions: Implications for fine-scale passive restoration. J. Ecol. Eng. 2019, 135, 28–35. [Google Scholar] [CrossRef]
- Feng, G.; Huang, J.; Xu, Y.; Li, J.; Zhang, R. Disentangling environmental effects on the tree species abundance distribution and richness in a subtropical forest. Front. Plant Sci. 2021, 12, 367. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.W. Leye Tiankeng; Guangxi Science and Technology Publishing House: Nanning, China, 2018. (In Chinese) [Google Scholar]
- Lin, Y. Species Diversity of Karst Tiankeng Forest in Dashiwei Tiankengs, Guangxi. Master Thesis, College of life Sciences, Guangxi Normal University, Guilin, China, 2005. (In Chinese). [Google Scholar]
- Pu, G.; Lv, Y.; Dong, L.; Zhou, L.; Huang, K.; Zeng, D.; Mo, L.; Xu, G. Profiling the bacterial diversity in a typical Karst Tiankeng of China. Biomolecules 2019, 9, 187. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.F.; Jiang, C.; Shui, W.; Guo, P.P.; Zhang, Y.Y.; Feng, J. Vertical distribution characteristics of plant community in shady slope of degraded Tiankeng talus: A case study of Zhanyi Shenxiantang in Yunnan, China. J. Appl. Ecol. 2020, 31, 1496–1504. [Google Scholar] [CrossRef]
- Hong, S.K.; Song, I.J.; Wu, J. Fengshui theory in urban landscape planning. Urban Ecosyst. 2007, 10, 221–237. [Google Scholar] [CrossRef]
- Su, Y.Q.; Tang, Q.M.; Mo, F.Y.; Xue, Y.G. Karst Tiankengs as refugia for indigenous tree flora amidst a degraded landscape in southwestern China. Sci. Rep. 2017, 7, 4249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Lin, W.; Zhang, Y.; Dai, Y.; Chen, B. Village Fengshui Forests as Forms of Cultural and Ecological Heritage: Interpretations and Conservation Policy Implications from Southern China. Forests 2020, 11, 1286. [Google Scholar] [CrossRef]
- Wan, F.; Zhang, X.; Li, T. Stability analysis of Jiguanshan Tunnel construction under karst tiankeng. IOP Conf. Ser. EES 2021, 769, 32–54. Available online: https://schlr.cnki.net/Detail/doi/GARJ2021_1/SIPD0DEE5CEC6C628C2AF275723D00351265 (accessed on 8 December 2021). [CrossRef]
- Huang, L.J.; Yu, Y.M.; An, X.F.; Yu, L.L.; Xue, Y.G. Interspecific association of main woody plants in Tiankeng forests of Dashiwei Tiankeng Group, Guangxi. Guihaia 2021, 41, 695–706. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, S.Z. Study on the Maintenance Mechanisms of Species Diversity in the Matural Old Growth Tropical Forests on the Hainan Island, China. Ph.D. Dissertation, Chinese Academy of Forestry, Beijing, China, 2017. (In Chinese). [Google Scholar]
- Wei, Y.; Wen, F.; Fu, L.; Xin, Z.; Jiang, R.; Pan, B.; Hua, F.; Mao, S.; Li, S.; Ma, H.; et al. The Distribution and Conservation Status of Native Plants in Guangxi, China; China Forestry Publishing: Beijing, China, 2018. [Google Scholar]
- Avolio, M.; Forrestel, E.J.; Chang, C.C.; La Pierre, K.J.; Burghardt, K.T.; Smith, M.D. Demystifying dominant species. New Phytol. 2019, 223, 1106–1126. [Google Scholar] [CrossRef] [PubMed]
- Skeen, J.N. An extension of the concept of importance value in analyzing forest communities. Ecology 1973, 54, 655–656. [Google Scholar] [CrossRef]
- Hadley, W.; Romain, F.; Lionel, H.; Kirill, M. Dplyr: A Grammar of Data Manipulation. R Package Version 1.0.0. 2020. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 9 December 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 8 December 2021).
- Keylock, C.J. Simpson diversity and the Shannon-wiener index as special cases of a generalized entropy. Oikos 2005, 109, 203–207. [Google Scholar] [CrossRef]
- Fletcher, S.; Islam, M. Comparing sets of patterns with the Jaccard index. Asian-Australas. J. Anim. Sci. 2018, 22, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Fisher, C.K.; Mehta, P. The transition between the niche and neutral regimes in ecology. Proc. Natl. Acad. Sci. USA 2014, 111, 13111–13116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, D.; Mckane, A.J. Sampling Hubbell’s neutral theory of biodiversity. Ecol. Lett. 2004, 7, 901–910. [Google Scholar] [CrossRef]
- Ramesh, T. Trends and Changes in Hydroclimatic Variable; Elsevier: Cambridge, MA, USA, 2018. [Google Scholar]
- Paulo, I.P.; Murilo, D.M.; Andre, C. Maximum Likelihood Models for Species Abundance Distributions. R Packages Version 0.4.2. 2018. Available online: http://piLaboratory.github.io/sads (accessed on 8 December 2021).
- Muluneh, M.; Feyissa, M.; Wolde, T. Effect of forest fragmentation and disturbance on diversity and structure of woody species in dry Afromontane forests of northern Ethiopia. Biodivers. Conserv. 2021, 30, 1753–1779. [Google Scholar] [CrossRef]
- Bertassello, L.; Bertuzzo, E.; Botter, G.; Jawitz, J.W.; Aubeneau, A.F.; Hoverman, J.T.; Rinaldo, A.; Rao, P.S.C. Dynamic spatio-temporal patterns of metapopulation occupancy in patchy habitats. R. Soc. Open. Sci. 2021, 8, 201309. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.X.; Coggins, C.; Minor, J.; Zhang, Y.Q. Fengshui forests and village landscapes in China: Geographic extent, socioecological significance, and conservation prospects. Urban For. Urban Green. 2018, 31, 79–92. [Google Scholar] [CrossRef]
- Shen, H.T.; Kimikazu, S.; Qi, M.; Masumi, M.; Tetsuya, M.; Seiji, H.; Takahashi, T.; Honda, M.; Sueki, K.; He, M.; et al. Exposure age dating of Chinese Tiankengs by 36Cl-AMS. Nucl. Instrum. Methods B 2019, 459, 29–35. [Google Scholar] [CrossRef]
- Yang, G.; Peng, C.; Liu, Y.; Dong, F.Q. Tiankeng: An ideal place for climate warming research on forest ecosystems. Environ. Earth Sci. 2019, 78, 46. [Google Scholar] [CrossRef]
- Wang, Y.; Lamontagne, J.M.; Lin, F.; Yuan, Z.Q.; Ye, J.; Wang, X.G.; Hao, Z. Similarity between seed rain and neighbouring mature tree communities in an old-growth temperate forest. J. For. Res. 2019, 31, 2435–2444. [Google Scholar] [CrossRef] [Green Version]
- Niccolo, A.; Jorge, H.; Carlos, A.P.; Tommaso, B.; Amos, M.; Samir, S. Neutral and niche forces as drivers of species selection. J. Theor. Biol. 2019, 483, 109969. [Google Scholar] [CrossRef]
- Pinsky, M.L. Species coexistence through competition and rapid evolution. Proc. Natl. Acad. Sci. USA 2019, 116, 2407–2409. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Species | The Inside of Tiankeng Forests | The Fringe of Tiankeng Forests | Nearby Non-Tiankeng Forests | |||
|---|---|---|---|---|---|---|
| DC-TK | LX-TK | LJ-BY | SM-BY | LA-FS | SW-FS | |
| Lindera glauca (Sieb. et Zucc.) Bl. | 29.82% | 9.27% | - | - | - | - |
| Schefflera guizhouensis C. B. Shang | 21.44% | 4.85% | - | - | - | - |
| Miliusa sinensis Finet et Gagnep. | 11.29% | - | - | - | - | - |
| Machilus chinensis (Champ. ex Benth.) Hemsl. | 10.69% | - | 3.88% | - | - | - |
| Handeliodendron bodinieri (Lévl.) Rehd. | 4.90% | 12.32% | 6.05% | 2.18% | 2.91% | 3.38% |
| Choerospondias axillaris (Roxb.) B. L. Burtt & A. W. Hill | 5.72% | 10.97% | 19.61% | - | 3.81% | 4.84% |
| Lindera pulcherrima var. Hemsleyana (Diels) H.P.Tsui | 1.29% | 9.90% | 5.78% | - | 2.97% | 1.10% |
| Ilex macrocarpa Oliv. | - | - | 10.83% | - | 4.79% | 9.99% |
| Itea macrophylla Wall. ex Roxb. | - | - | 10.05% | - | - | - |
| Machilus glaucifolia S. K. Lee & F. N. Wei | - | - | 8.97% | 15.39% | - | - |
| Celtis sinensis Pers. | 1.10% | 8.61% | 2.67% | - | 11.70% | 7.84% |
| Rhaphiolepis indica (Linnaeus) Lindley | - | 1.73% | 1.64% | - | 3.15% | 12.54% |
| Illicium simonsii Maxim. | - | - | - | - | 10.82% | - |
| Cinnamomum bodinieri Lévl. | - | - | 2.47% | - | 1.05% | 10.45% |
| Litsea rotundifolia var. Oblongifolia (Nees) Allen | - | 1.62% | 9.78% | - | 1.26% | - |
| Itea yunnanensis Franch. | - | 8.20% | 8.82% | - | 5.83% | 9.61% |
| Neutral Theory Parameter | Species Diversity Index | ||||
|---|---|---|---|---|---|
| Volkov Model | Zero-Sum Polynomials Model | ||||
| Sites | θ | m | θ | Species richness | Shannon-Wiener index |
| DC-TK | 8.31 | 0.35 | 6.71 | 31 | 3.81 |
| LX-TK | 7.3 | 0.4 | 7.24 | 32 | 3.64 |
| LJ-BY | 16.65 | 0.59 | 14.28 | 48 | 4.63 |
| SM-BY | 15.34 | 0.62 | 12.44 | 50 | 4.40 |
| LA-FS | 13.34 | 0.41 | 11.12 | 47 | 4.33 |
| SW-FS | 14.75 | 0.42 | 11.86 | 41 | 4.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, L.; Yang, H.; An, X.; Yu, Y.; Yu, L.; Huang, G.; Liu, X.; Chen, M.; Xue, Y. Species Abundance Distributions Patterns between Tiankeng Forests and Nearby Non-Tiankeng Forests in Southwest China. Diversity 2022, 14, 64. https://doi.org/10.3390/d14020064
Huang L, Yang H, An X, Yu Y, Yu L, Huang G, Liu X, Chen M, Xue Y. Species Abundance Distributions Patterns between Tiankeng Forests and Nearby Non-Tiankeng Forests in Southwest China. Diversity. 2022; 14(2):64. https://doi.org/10.3390/d14020064
Chicago/Turabian StyleHuang, Linjuan, Hao Yang, Xiaofei An, Yanmei Yu, Linlan Yu, Gui Huang, Xinyu Liu, Ming Chen, and Yuegui Xue. 2022. "Species Abundance Distributions Patterns between Tiankeng Forests and Nearby Non-Tiankeng Forests in Southwest China" Diversity 14, no. 2: 64. https://doi.org/10.3390/d14020064
APA StyleHuang, L., Yang, H., An, X., Yu, Y., Yu, L., Huang, G., Liu, X., Chen, M., & Xue, Y. (2022). Species Abundance Distributions Patterns between Tiankeng Forests and Nearby Non-Tiankeng Forests in Southwest China. Diversity, 14(2), 64. https://doi.org/10.3390/d14020064