
Neurosurgical Approaches to Brain Tissue Harvesting for the Establishment of Cell Cultures in Neural Experimental Cell Models



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Sources for Brain Cell Isolation
3. The Challenges of Isolating the Brain Tissue Cells
4. A Brief Description for a Culture Protocol for Brain Cells
4.1. The Source of the Tissue
4.2. Preparation of Tissue for Cell Culture
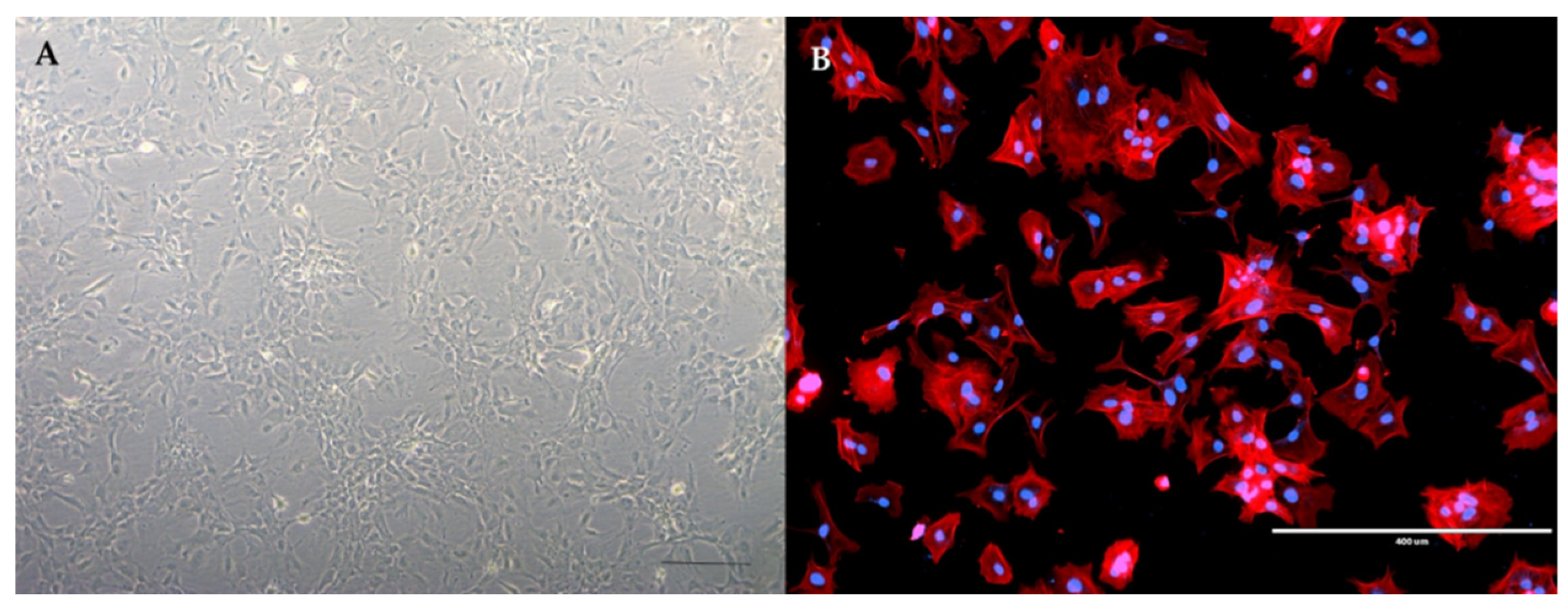
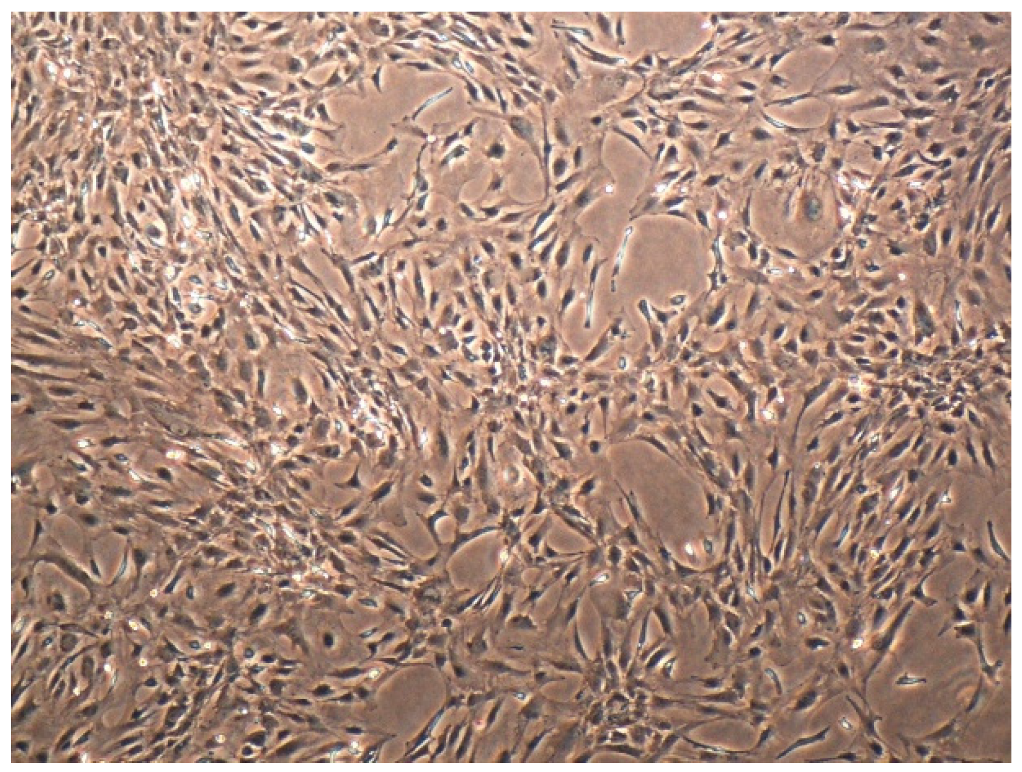
4.3. The Culture of Primary Astrocytes
5. Brain Cells and Biomaterial Development
6. Relationships between Tissue Sampling and Biomaterial Testing
7. Neurosurgical Approaches for Brain Tissue Sampling
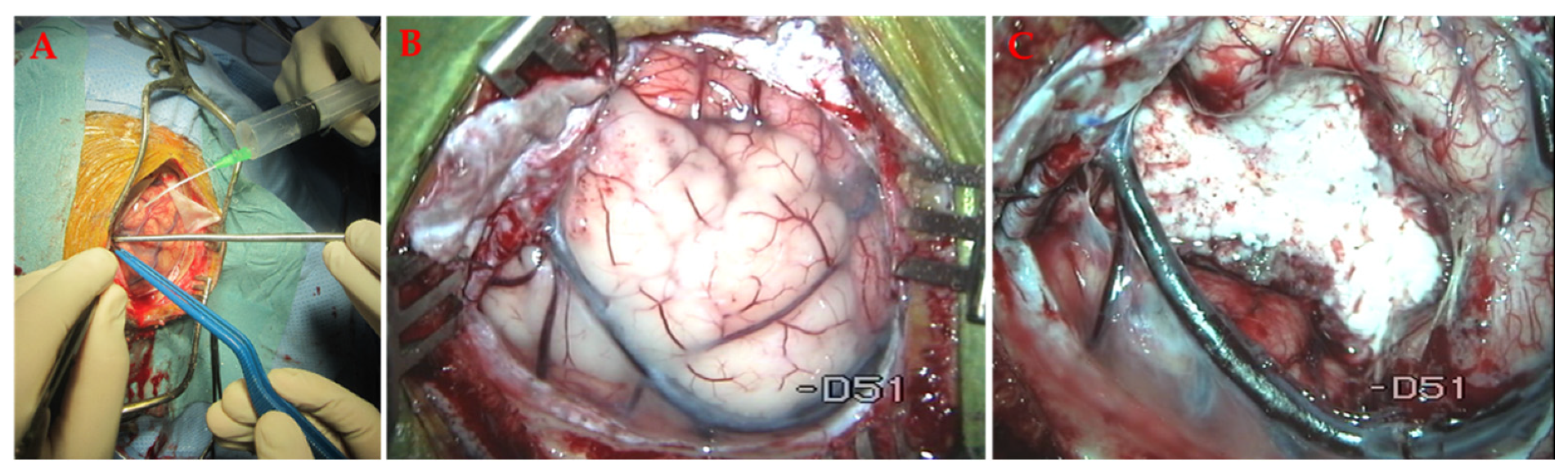
7.1. Open Surgery
7.2. Keyhole Approaches
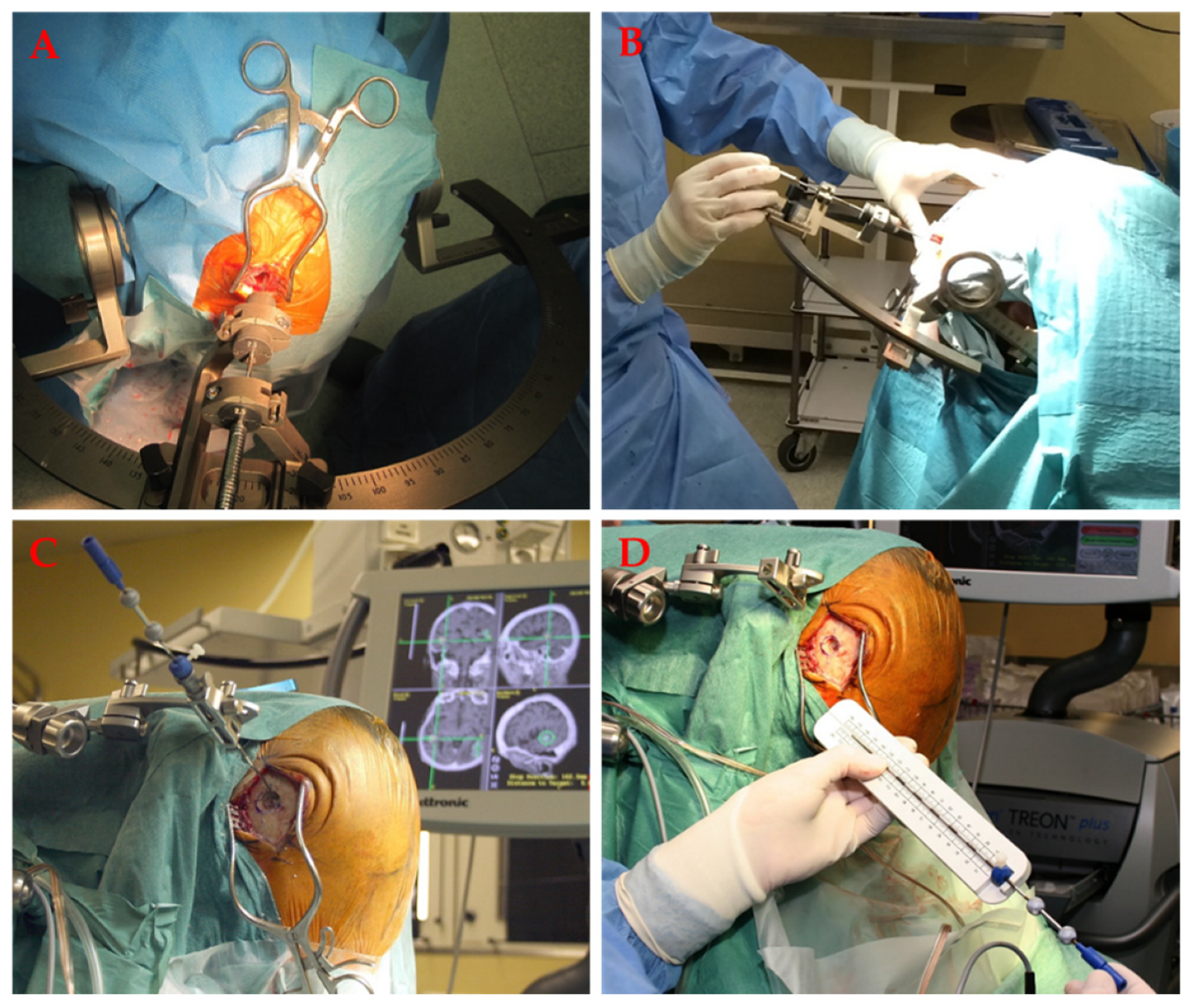
7.3. Stereotactic Needle Biopsy
7.4. Neuroendoport Surgery
7.5. Neuroendoscopic Surgery
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, A.; Madden, L.A.; Paunov, V.N. Advanced biomedical applications based on emerging 3D cell culturing platforms. J. Mater. Chem. B 2020, 8, 10487–10501. [Google Scholar] [CrossRef]
- Keller, J.M.; Frega, M. Past, present, and future of neuronal models in vitro. In Vitro Neuronal Networks; Springer Nature: Basingstoke, UK, 2019; pp. 3–17. [Google Scholar]
- Bassi, G.; Grimaudo, M.A.; Panseri, S.; Montesi, M. Advanced multi-dimensional cellular models as emerging reality to reproduce In Vitro the human body complexity. Int. J. Mol. Sci. 2021, 22, 1195. [Google Scholar] [CrossRef]
- Trope, C.; Sigurdsson, K. Use of tissue culture in predictive testing of drug sensitivity in human ovarian cancer. Correlation between in vitro results and the response in vivo. Neoplasma 1982, 29, 309–314. [Google Scholar]
- Bermudez-Brito, M.; Plaza-Diaz, J.; Fontana, L.; Munoz-Quezada, S.; Gil, A. In vitro cell and tissue models for studying host-microbe interactions: A review. Br. J. Nutr. 2013, 109, S27–S34. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Xiang, S.; Li, E.N.; Fritch, M.R.; Alexander, P.G.; Lin, H.; Tuan, R.S. Tissue Engineering for Musculoskeletal Regeneration and Disease Modeling; Springer Nature: Basingstoke, UK, 2020; pp. 235–268. [Google Scholar]
- Freshney, R.I. Basic Principles of Cell Culture; John Wiley & Sons, Inc.: Hobokan, NJ, USA, 2006. [Google Scholar]
- Mather, J.P. In vitro models. Stem Cells 2012, 30, 95–99. [Google Scholar] [CrossRef]
- Freshney, R. Culture of Animal Cells: A Manual of Basic Technique. Chapter 2: Biology of Cultured Cells; Willey-Liss. Inc.: New York, NY, USA, 2000; pp. 9–15. [Google Scholar]
- Francis, S.L.; Yao, A.; Choong, P.F. Culture Time Needed to Scale up Infrapatellar Fat Pad Derived Stem Cells for Cartilage Regeneration: A Systematic Review. Bioengineering 2020, 7, 69. [Google Scholar] [CrossRef]
- Kobayashi, J.; Kikuchi, A.; Aoyagi, T.; Okano, T. Cell sheet tissue engineering: Cell sheet preparation, harvesting/manipulation, and transplantation. J. Biomed. Mater. Res. A 2019, 107, 955–967. [Google Scholar] [CrossRef]
- Centeno, C.J. Clinical challenges and opportunities of mesenchymal stem cells in musculoskeletal medicine. PM&R 2014, 6, 70–77. [Google Scholar]
- Knowlton, S.; Anand, S.; Shah, T.; Tasoglu, S. Bioprinting for neural tissue engineering. Trends Neurosci. 2018, 41, 31–46. [Google Scholar] [CrossRef]
- Azzarelli, R. Organoid models of glioblastoma to study brain tumor stem cells. Front. Cell Dev. Biol. 2020, 8, 220. [Google Scholar] [CrossRef] [Green Version]
- DiSano, K.D.; Linzey, M.R.; Welsh, N.C.; Meier, J.S.; Pachner, A.R.; Gilli, F. Isolating Central Nervous System Tissues and Associated Meninges for the Downstream Analysis of Immune cells. JoVE J. Vis. Exp. 2020, 159, e61166. [Google Scholar] [CrossRef]
- Zhu, Z.; Cui, W.; Zhu, D.; Gao, N.; Zhu, Y. Common tools for pituitary adenomas research: Cell lines and primary cells. Pituitary 2020, 23, 182–188. [Google Scholar] [CrossRef]
- Gradisnik, L.; Maver, U.; Bosnjak, R.; Velnar, T. Optimised isolation and characterisation of adult human astrocytes from neurotrauma patients. J. Neurosci. Methods 2020, 341, 108796. [Google Scholar] [CrossRef]
- Chesnelong, C.; Restall, I.; Weiss, S. Isolation and culture of glioblastoma brain tumor stem cells. In Brain Tumor Stem Cells; Springer: Berlin, Germany, 2019; pp. 11–21. [Google Scholar]
- Rustenhoven, J.; Park, T.I.; Schweder, P.; Scotter, J.; Correia, J.; Smith, A.M.; Gibbons, H.M.; Oldfield, R.L.; Bergin, P.S.; Mee, E.W. Isolation of highly enriched primary human microglia for functional studies. Sci. Rep. 2016, 6, 19371. [Google Scholar] [CrossRef] [Green Version]
- Tanti, G.K.; Srivastava, R.; Kalluri, S.R.; Nowak, C.; Hemmer, B. Isolation, Culture and Functional Characterization of Glia and Endothelial Cells from Adult Pig Brain. Front. Cell. Neurosci. 2019, 13, 333. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, C.E.; Alvendia, H.; Halliday, G.M. Brain banking for research into neurodegenerative disorders and ageing. Neurosci. Bull. 2019, 35, 283–288. [Google Scholar] [CrossRef]
- Huszthy, P.C.; Daphu, I.; Niclou, S.P.; Stieber, D.; Nigro, J.M.; Sakariassen, P.Ø.; Miletic, H.; Thorsen, F.; Bjerkvig, R. In vivo models of primary brain tumors: Pitfalls and perspectives. Neuro-Oncology 2012, 14, 979–993. [Google Scholar] [CrossRef] [Green Version]
- Venkataraman, L.; Fair, S.R.; McElroy, C.A.; Hester, M.E.; Fu, H. Modeling neurodegenerative diseases with cerebral organoids and other three-dimensional culture systems: Focus on Alzheimer’s disease. Stem Cell Rev. Rep. 2020. [Google Scholar] [CrossRef]
- Hojat, A.; Wei, B.; Olson, M.G.; Mao, Q.; Yong, W.H. Procurement and storage of surgical biospecimens. Biobanking 2019. [Google Scholar] [CrossRef]
- Zielke, H.R.; Mash, D.C. A review of brain biorepository management and operations. Handb. Clin. Neurol. 2018, 150, 83–92. [Google Scholar]
- Chew, L.-J.; DeBoy, C.A.; Senatorov, V.V., Jr. Finding degrees of separation: Experimental approaches for astroglial and oligodendroglial cell isolation and genetic targeting. J. Neurosci. Methods 2014, 236, 125–147. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denis-Donini, S.; Glowinski, J.; Prochiantz, A. Glial heterogeneity may define the three-dimensional shape of mouse mesencephalic dopaminergic neurones. Nature 1984, 307, 641–643. [Google Scholar] [CrossRef]
- Garcia-Abreu, J.; Neto, V.M.; Carvalho, S.; Cavalcante, L. Regionally specific properties of midbrain glia: I. Interactions with midbrain neurons. J. Neurosci. Res. 1995, 40, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Kimelberg, H.K. The problem of astrocyte identity. Neurochem. Int. 2004, 45, 191–202. [Google Scholar] [CrossRef]
- Lee, S.C.; Liu, W.; Brosnan, C.F.; Dickson, D.W. Characterization of primary human fetal dissociated central nervous system cultures with an emphasis on microglia. Lab. Investig. J. Tech. Methods Pathol. 1992, 67, 465–476. [Google Scholar]
- Sharif, A.; Prevot, V. Isolation and culture of human astrocytes. In Astrocytes; Springer: Berlin, Germany, 2012; pp. 137–151. [Google Scholar]
- Oberheim, N.A.; Takano, T.; Han, X.; He, W.; Lin, J.H.; Wang, F.; Xu, Q.; Wyatt, J.D.; Pilcher, W.; Ojemann, J.G. Uniquely hominid features of adult human astrocytes. J. Neurosci. 2009, 29, 3276–3287. [Google Scholar] [CrossRef]
- Chaboub, L.S.; Deneen, B. Developmental origins of astrocyte heterogeneity: The final frontier of CNS development. Dev. Neurosci. 2012, 34, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Bignami, A.; Eng, L.; Dahl, D.; Uyeda, C. Localization of the glial fibrillary acidic protein in astrocytes by immunofluorescence. Brain Res. 1972, 43, 429–435. [Google Scholar] [CrossRef]
- Giffard, R.; Ouyang, Y.-B. Cell Culture: Primary Neural Cells; Elsevier: London, UK, 2009. [Google Scholar]
- Nakagawa, T.; Schwartz, J.P. Gene expression patterns in in vivo normal adult astrocytes compared with cultured neonatal and normal adult astrocytes. Neurochem. Int. 2004, 45, 203–242. [Google Scholar] [CrossRef]
- John, G.R. Investigation of astrocyte–oligodendrocyte interactions in human cultures. In Astrocytes; Springer: Berlin, Germany, 2012; pp. 401–414. [Google Scholar]
- Sharif, A.; Prevot, V.; Renault-Mihara, F.; Allet, C.; Studler, J.; Canton, B.; Chneiweiss, H.; Junier, M. Transforming growth factor alpha acts as a gliatrophin for mouse and human astrocytes. Oncogene 2006, 25, 4076–4085. [Google Scholar] [CrossRef] [Green Version]
- Minchev, G.; Kronreif, G.; Ptacek, W.; Dorfer, C.; Micko, A.; Maschke, S.; Legnani, F.G.; Widhalm, G.; Knosp, E.; Wolfsberger, S. A novel robot-guided minimally invasive technique for brain tumor biopsies. J. Neurosurg. 2019, 132, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Dammers, R.; Haitsma, I.; Schouten, J.; Kros, J.; Avezaat, C.; Vincent, A. Safety and efficacy of frameless and frame-based intracranial biopsy techniques. Acta Neurochir. 2008, 150, 23–29. [Google Scholar] [CrossRef]
- Middeldorp, J.; Hol, E. GFAP in health and disease. Prog. Neurobiol. 2011, 93, 421–443. [Google Scholar] [CrossRef]
- Barres, B.A. Designing and troubleshooting immunopanning protocols for purifying neural cells. Cold Spring Harb. Protoc. 2014, 2014, 1342–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, D. Astrocytes: Form, functions, and roles in disease. Vet. Pathol. 1994, 31, 145–167. [Google Scholar] [CrossRef]
- Jakovcevski, I.; Filipovic, R.; Mo, Z.; Rakic, S.; Zecevic, N. Oligodendrocyte development and the onset of myelination in the human fetal brain. Front. Neuroanat. 2009, 3, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, S.A.; Kuypers, N.J. How to make an oligodendrocyte. Development 2015, 142, 3983–3995. [Google Scholar] [CrossRef] [Green Version]
- Sarthy, V. Focus on molecules: Glial fibrillary acidic protein (GFAP). Exp. Eye Res. 2006, 84, 381–382. [Google Scholar] [CrossRef]
- Dorfer, C.; Rydenhag, B.; Baltuch, G.; Buch, V.; Blount, J.; Bollo, R.; Gerrard, J.; Nilsson, D.; Roessler, K.; Rutka, J. How technology is driving the landscape of epilepsy surgery. Epilepsia 2020, 61, 841–855. [Google Scholar] [CrossRef] [PubMed]
- Micera, S.; Caleo, M.; Chisari, C.; Hummel, F.C.; Pedrocchi, A. Advanced neurotechnologies for the restoration of motor function. Neuron 2020, 105, 604–620. [Google Scholar] [CrossRef]
- Kimelberg, H.K.; Schools, G.P.; Cai, Z.; Zhou, M. Freshly isolated astrocyte (FIA) preparations: A useful single cell system for studying astrocyte properties. J. Neurosci. Res. 2000, 61, 577–587. [Google Scholar] [CrossRef]
- Velnar, T.; Maver, U.; Bosnjak, R.; Gradisnik, L. The Isolation of Human Glioblastoma Cells: An Optimised Protocol. Acta Med. Acad. 2020, 49, 4–13. [Google Scholar] [CrossRef]
- Dundar, T.T.; Hatiboglu, M.A.; Ergul, Z.; Seyithanoglu, M.H.; Sozen, E.; Tuzgen, S.; Kaynar, M.Y.; Karaoz, E. Glioblastoma stem cells and comparison of isolation methods. J. Clin. Med. Res. 2019, 11, 415. [Google Scholar] [CrossRef] [Green Version]
- Wu, V.W.; Nishiyama, N.; Schwartz, J.P. A culture model of reactive astrocytes: Increased nerve growth factor synthesis and reexpression of cytokine responsiveness. J. Neurochem. 1998, 71, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Mignot, C.; Delarasse, C.; Escaich, S.; Della Gaspera, B.; Noé, E.; Colucci-Guyon, E.; Babinet, C.; Pekny, M.; Vicart, P.; Boespflug-Tanguy, O. Dynamics of mutated GFAP aggregates revealed by real-time imaging of an astrocyte model of Alexander disease. Exp. Cell Res. 2007, 313, 2766–2779. [Google Scholar] [CrossRef]
- Mizee, M.R.; Miedema, S.S.; van der Poel, M.; Adelia; Schuurman, K.G.; van Strien, M.E.; Melief, J.; Smolders, J.; Hendrickx, D.A.; Heutinck, K.M.; et al. Isolation of primary microglia from the human post-mortem brain: Effects of ante- and post-mortem variables. Acta Neuropathol. Commun. 2017, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimmerjahn, A. Astrocytes going live: Advances and challenges. J. Physiol. 2009, 587, 1639–1647. [Google Scholar] [CrossRef]
- Zhang, Y.; Barres, B.A. Astrocyte heterogeneity: An underappreciated topic in neurobiology. Curr. Opin. Neurobiol. 2010, 20, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Jungblut, M.; Tiveron, M.C.; Barral, S.; Abrahamsen, B.; Knöbel, S.; Pennartz, S.; Schmitz, J.; Perraut, M.; Pfrieger, F.W.; Stoffel, W. Isolation and characterization of living primary astroglial cells using the new GLAST-specific monoclonal antibody ACSA-1. Glia 2012, 60, 894–907. [Google Scholar] [CrossRef]
- Levine, J.M. Neuronal influences on glial progenitor cell development. Neuron 1989, 3, 103–113. [Google Scholar] [CrossRef]
- Lange, S.C.; Bak, L.K.; Waagepetersen, H.S.; Schousboe, A.; Norenberg, M.D. Primary Cultures of Astrocytes: Their Value in Understanding Astrocytes in Health and Disease. Neurochem. Res. 2012, 37, 2569–2588. [Google Scholar] [CrossRef] [Green Version]
- Parkhurst, C.N.; Gan, W.-B. Microglia dynamics and function in the CNS. Curr. Opin. Neurobiol. 2010, 20, 595–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Condic, M.; Oberstein, T.J.; Herrmann, M.; Reimann, M.C.; Kornhuber, J.; Maler, J.M.; Spitzer, P. N-truncation and pyroglutaminylation enhances the opsonizing capacity of Aβ-peptides and facilitates phagocytosis by macrophages and microglia. Brain Behav. Immun. 2014, 41, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Agalave, N.M.; Lane, B.T.; Mody, P.H.; Szabo-Pardi, T.A.; Burton, M.D. Isolation, culture, and downstream characterization of primary microglia and astrocytes from adult rodent brain and spinal cord. J. Neurosci. Methods 2020, 340, 108742. [Google Scholar] [CrossRef]
- Uliasz, T.F.; Hamby, M.E.; Jackman, N.A.; Hewett, J.A.; Hewett, S.J. Generation of primary astrocyte cultures devoid of contaminating microglia. In Astrocytes; Springer: Berlin, Germany, 2012; pp. 61–79. [Google Scholar]
- Welser, J.V.; Milner, R. Derivation of Microglia-Free Astrocyte Cultures from Neural Stem Cells. In Astrocytes; Springer: Berlin, Germany, 2012; pp. 81–91. [Google Scholar]
- Wolf, F.; Kirchhoff, F. Neuroscience. Imaging astrocyte activity. Science 2008, 320, 1597–1599. [Google Scholar] [CrossRef] [PubMed]
- Araque, A.; Sanzgiri, R.P.; Parpura, V.; Haydon, P.G. Astrocyte-induced modulation of synaptic transmission. Can. J. Physiol. Pharmacol. 1999, 77, 699–706. [Google Scholar] [CrossRef]
- Sofroniew, M.V. Reactive astrocytes in neural repair and protection. Neuroscientist 2005, 11, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Berishvili, E.; Casiraghi, F.; Amarelli, C.; Scholz, H.; Piemonti, L.; Berney, T.; Montserrat, N. Mini-organs forum: How to advance organoid technology to organ transplant community. Transpl. Int. 2021, 34, 1588–1593. [Google Scholar] [CrossRef]
- Khaseb, S.; Orooji, M.; Pour, M.G.; Safavi, S.M.; Eghbal, M.J.; Rad, M.R. Dental stem cell banking: Techniques and Protocols. Cell Biol. Int. 2021, 45, 1–15. [Google Scholar] [CrossRef]
- Lee, S.-G.; Su, Z.-Z.; Emdad, L.; Gupta, P.; Sarkar, D.; Borjabad, A.; Volsky, D.J.; Fisher, P.B. Mechanism of ceftriaxone induction of excitatory amino acid transporter-2 expression and glutamate uptake in primary human astrocytes. J. Biol. Chem. 2008, 283, 13116–13123. [Google Scholar] [CrossRef] [Green Version]
- Caffrey, T.M.; Button, E.B.; Robert, J. Toward three-dimensional in vitro models to study neurovascular unit functions in health and disease. Neural Regen. Res. 2021, 16, 2132. [Google Scholar]
- Lecrux, C.; Hamel, E. The neurovascular unit in brain function and disease. Acta Physiol. 2011, 203, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Haggerty, A.E.; Maldonado-Lasunción, I.; Oudega, M. Biomaterials for revascularization and immunomodulation after spinal cord injury. Biomed. Mater. 2018, 13, 044105. [Google Scholar] [CrossRef] [Green Version]
- Oudega, M. Molecular and cellular mechanisms underlying the role of blood vessels in spinal cord injury and repair. Cell Tissue Res. 2012, 349, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Straley, K.S.; Foo, C.W.P.; Heilshorn, S.C. Biomaterial design strategies for the treatment of spinal cord injuries. J. Neurotrauma 2010, 27, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Macaya, D.; Spector, M. Injectable hydrogel materials for spinal cord regeneration: A review. Biomed. Mater. 2012, 7, 012001. [Google Scholar] [CrossRef]
- Pakulska, M.M.; Ballios, B.G.; Shoichet, M.S. Injectable hydrogels for central nervous system therapy. Biomed. Mater. 2012, 7, 024101. [Google Scholar] [CrossRef]
- Kimelberg, H.K.; Nedergaard, M. Functions of astrocytes and their potential as therapeutic targets. Neurotherapeutics 2010, 7, 338–353. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Yang, Z.; Li, X. Regeneration strategies after the adult mammalian central nervous system injury—Biomaterials. Regen. Biomater. 2016, 3, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Cregg, J.M.; DePaul, M.A.; Filous, A.R.; Lang, B.T.; Tran, A.; Silver, J. Functional regeneration beyond the glial scar. Exp. Neurol. 2014, 253, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Montani, L.; Petrinovic, M.M. Targeting axonal regeneration: The growth cone takes the lead. J. Neurosci. 2014, 34, 4443–4444. [Google Scholar] [CrossRef] [Green Version]
- Struzyna, L.A.; Harris, J.P.; Katiyar, K.S.; Chen, H.I.; Cullen, D.K. Restoring nervous system structure and function using tissue engineered living scaffolds. Neural Regen. Res. 2015, 10, 679–685. [Google Scholar]
- Placone, A.L.; McGuiggan, P.M.; Bergles, D.E.; Guerrero-Cazares, H.; Quiñones-Hinojosa, A.; Searson, P.C. Human astrocytes develop physiological morphology and remain quiescent in a novel 3D matrix. Biomaterials 2015, 42, 134–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möllers, S.; Heschel, I.; Damink, L.H.O.; Schügner, F.; Deumens, R.; Müller, B.; Bozkurt, A.; Nava, J.G.; Noth, J.; Brook, G.A. Cytocompatibility of a novel, longitudinally microstructured collagen scaffold intended for nerve tissue repair. Tissue Eng. Part A 2009, 15, 461–472. [Google Scholar] [CrossRef]
- Lie, D.C.; Song, H.; Colamarino, S.A.; Ming, G.-l.; Gage, F.H. Neurogenesis in the adult brain: New strategies for central nervous system diseases. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 399–421. [Google Scholar] [CrossRef] [Green Version]
- Toy, D.; Namgung, U. Role of glial cells in axonal regeneration. Exp. Neurobiol. 2013, 22, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Courtney, J.; Irvine, L.; Jones, C.; Mosa, S.; Robertson, L.; Srivastava, S. Biomaterials in medicine-a bioengineering perspective. Int. J. Artif. Organs 1993, 16, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M.; Gristina, A.G.; Hanson, S.R.; Harker, L.A.; Johnson, R.J.; Merritt, K.; Naylor, P.T.; Schoen, F.J. Host reactions to biomaterials and their evaluation. In Biomaterials Science; Elsevier: Amsterdam, The Netherlands, 1996; pp. 165–214. [Google Scholar]
- Greisler, H.P.; Dennis, J.W.; Endean, E.D.; Ellinger, J.; Buttle, K.F.; Kim, D.U. Derivation of neointima in vascular grafts. Circulation 1988, 78, I6–I12. [Google Scholar] [PubMed]
- Lindenauer, S.; Stanley, J.; Zelenock, G.; Cronenwett, J.; Whitehouse, W., Jr.; Erlandson, E. Aorto-iliac reconstruction with Dacron double velour. J. Cardiovasc. Surg. 1984, 25, 36–42. [Google Scholar]
- Wilkinson, C. Making structures for cell engineering. Eur. Cells Mater. 2004, 8, 21–26. [Google Scholar] [CrossRef]
- Sigler, M.; Paul, T.; Grabitz, R. Biocompatibility screening in cardiovascular implants. Z. Fur Kardiol. 2005, 94, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Greisler, H.P.; Petsikas, D.; Lam, T.M.; Patel, N.; Ellinger, J.; Cabusao, E.; Tattersall, C.W.; Kim, D.U. Kinetics of cell proliferation as a function of vascular graft material. J. Biomed. Mater. Res. 1993, 27, 955–961. [Google Scholar] [CrossRef]
- Padera, R.F.; Colton, C.K. Time course of membrane microarchitecture-driven neovascularization. Biomaterials 1996, 17, 277–284. [Google Scholar] [CrossRef]
- Kirkpatrick, C.; Krump-Konvalinkova, V.; Unger, R.; Bittinger, F.; Otto, M.; Peters, K. Tissue response and biomaterial integration: The efficacy of in vitro methods. Biomol. Eng. 2002, 19, 211–217. [Google Scholar] [CrossRef]
- Tiwari, A.; Salacinski, H.; Seifalian, A.M.; Hamilton, G. New prostheses for use in bypass grafts with special emphasis on polyurethanes. Cardiovasc. Surg. 2002, 10, 191–197. [Google Scholar] [CrossRef]
- Brown, B.N.; Badylak, S.F. Expanded applications, shifting paradigms and an improved understanding of host–biomaterial interactions. Acta Biomater. 2013, 9, 4948–4955. [Google Scholar] [CrossRef]
- Brown, B.N.; Londono, R.; Tottey, S.; Zhang, L.; Kukla, K.A.; Wolf, M.T.; Daly, K.A.; Reing, J.E.; Badylak, S.F. Macrophage phenotype as a predictor of constructive remodeling following the implantation of biologically derived surgical mesh materials. Acta Biomater. 2012, 8, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Trandafir, A.F.; Dorin Eugen, P.; Vasile, D. Prostheses used in laparoscopic inguinal hernia repair: Biocompatibility, postoperative complications and quality of life—Review of the literature. Maedica 2017, 12, 202–207. [Google Scholar]
- Danaila, L.; Radoi, M. Surgery of tumors of the third ventricle region. Chirurgia 2013, 108, 456–462. [Google Scholar]
- Sudhakar, V.; Richardson, R.M. Gene therapy for neurodegenerative diseases. Neurotherapeutics 2019, 16, 166–175. [Google Scholar] [CrossRef] [Green Version]
- Savu, R.; Mohan, D.; Mohan, A.; Moisa, H.; Ciurea, A. A modern minimally invasive transcallosal approach to the third ventricle: A personal experience of 30 patients. Chirurgia 2014, 109, 80–89. [Google Scholar]
- Teo, C. The concept of minimally invasive neurosurgery. Neurosurg. Clin. N. Am. 2010, 21, 583–584. [Google Scholar] [CrossRef]
- Tatter, S.B. Neurosurgical management of brain tumors. Neuroimag. Clin. N. Am. 1999, 9, 779–799. [Google Scholar]
- Asthagiri, A.R.; Pouratian, N.; Sherman, J.; Ahmed, G.; Shaffrey, M.E. Advances in brain tumor surgery. Neurol. Clin. 2007, 25, 975–1003. [Google Scholar] [CrossRef] [PubMed]
- Tatter, S.B. Neurosurgical management of low-and intermediate-grade gliomas. Semin. Radiat. Oncolo. 2001, 11, 113–123. [Google Scholar] [CrossRef]
- Lacroix, M.; Abi-Said, D.; Fourney, D.R.; Gokaslan, Z.L.; Shi, W.; DeMonte, F.; Lang, F.F.; McCutcheon, I.E.; Hassenbusch, S.J.; Holland, E. A multivariate analysis of 416 patients with glioblastoma multiforme: Prognosis, extent of resection, and survival. J. Neurosurg. 2001, 95, 190–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamorano, L.; Kadi, A.; Dong, A. Computer-assisted neurosurgery: Simulation and automation. Stereot. Funct. Neurosurg. 1992, 59, 115–122. [Google Scholar] [CrossRef]
- Schaller, C.; Meyer, B.; Van Roost, D.; Schramm, J. Image guided microsurgery with a semifreehand neuronavigational device. Comput. Aided Surg. Off. J. Int. Soc. Comput. Aided Surg. ISCAS 1997, 2, 162–171. [Google Scholar] [CrossRef]
- Save, A.V.; Gill, B.J.; D’amico, R.S.; Canoll, P.; Bruce, J.N. Fluorescein-guided resection of gliomas. J. Neurosurg. Sci. 2019, 63, 648–655. [Google Scholar] [CrossRef]
- Hall, W.A. The safety and efficacy of stereotactic biopsy for intracranial lesions. Cancer Interdiscip. Int. J. Am. Cancer Soc. 1998, 82, 1749–1755. [Google Scholar] [CrossRef]
- Kvint, S.; Gutierrez, A.; Blue, R.; Petrov, D. Surgical Management of Trauma-Related Intracranial Hemorrhage—A Review. Curr. Neurol. Neurosci. Rep. 2020, 20, 63. [Google Scholar] [CrossRef]
- Wang, J.L.; Elder, J.B. Techniques for Open Surgical Resection of Brain Metastases. Neurosurg. Clin. 2020, 31, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D. Minor, moderate and severe head injury. Neurosurg. Rev. 1986, 9, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Recinos, P.F.; Raza, S.M.; Jallo, G.I.; Recinos, V.R. Use of a minimally invasive tubular retraction system for deep-seated tumors in pediatric patients. J. Neurosurg. Pediatr. 2011, 7, 516–521. [Google Scholar] [CrossRef] [Green Version]
- Rosenørn, J.; Diemer, N.H. Reduction of regional cerebral blood flow during brain retraction pressure in the rat. J. Neurosurg. 1982, 56, 826–829. [Google Scholar] [CrossRef]
- Miller, J.D. Head injury and brain ischaemia-implications for therapy. Br. J. Anaesth. 1985, 57, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Ciric, I. Aggressive surgery for malignant supratentorial gliomas. Clin. Neurosurg. 1990, 36, 375–383. [Google Scholar]
- Fadul, C.; Wood, J.; Thaler, H.; Galicich, J.; Patterson, R.; Posner, J. Morbidity and mortality of craniotomy for excision of supratentorial gliomas. Neurology 1988, 38, 1374. [Google Scholar] [CrossRef]
- Reisch, R.; Perneczky, A.; Filippi, R. Surgical technique of the supraorbital key-hole craniotomy. Surg. Neurol. 2003, 59, 223–227. [Google Scholar] [CrossRef]
- Iacoangeli, M.; Nocchi, N.; Nasi, D.; Di Rienzo, A.; Dobran, M.; Gladi, M.; Colasanti, R.; Alvaro, L.; Polonara, G.; Scerrati, M. Minimally invasive supraorbital key-hole approach for the treatment of anterior cranial fossa meningiomas. Neurol. Med.-Chir. 2016, 56, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Banu, M.A.; Mehta, A.; Ottenhausen, M.; Fraser, J.F.; Patel, K.S.; Szentirmai, O.; Anand, V.K.; Tsiouris, A.J.; Schwartz, T.H. Endoscope-assisted endonasal versus supraorbital keyhole resection of olfactory groove meningiomas: Comparison and combination of 2 minimally invasive approaches. J. Neurosurg. 2016, 124, 605–620. [Google Scholar] [CrossRef] [Green Version]
- Renfrow, J.J.; Strowd, R.E.; Laxton, A.W.; Tatter, S.B.; Geer, C.P.; Lesser, G.J. Surgical considerations in the optimal management of patients with malignant brain tumors. Curr. Treat. Options Oncol. 2017, 18, 46. [Google Scholar] [CrossRef] [PubMed]
- Reisch, R.; Stadie, A.; Kockro, R.; Gawish, I.; Schwandt, E.; Hopf, N. The minimally invasive supraorbital subfrontal key-hole approach for surgical treatment of temporomesial lesions of the dominant hemisphere. Min-Minim. Invasive Neurosurg. 2009, 52, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Gildenberg, P.L. The birth of human stereotactic surgery. In Stereot Funct Neuros; Springer: Berlin, Germany, 2013; pp. 1–4. [Google Scholar]
- Nashold, B., Jr. Stereotactic neurosurgery: The present and future. Am. Surg. 1970, 36, 91–93. [Google Scholar] [PubMed]
- Gildenberg, P.L.; Labuz, J. Use of a volumetric target for image-guided surgery. Neurosurgery 2006, 59, 651–659. [Google Scholar] [CrossRef]
- Bullard, D.E.; Nashold, B.S., Jr. Evolution of principles of stereotactic neurosurgery. Neurosurg. Clin. N. Am. 1995, 6, 27–41. [Google Scholar] [CrossRef]
- Maciunas, R.J.; Galloway, R.L., Jr.; Latimer, J.W. The application accuracy of stereotactic frames. Neurosurgery 1994, 35, 682–695. [Google Scholar] [CrossRef]
- Maciunas, R.J.; Galloway, R.L., Jr.; Latimer, J.; Cobb, C.; Zaccharias, E.; Moore, A.; Mandava, V.R. An independent application accuracy evaluation of stereotactic frame systems. Stereotact. Funct. Neurosurg. 1992, 58, 103–107. [Google Scholar] [CrossRef]
- Heilbrun, M.P.; McDonald, P.; Wiker, C.; Koehler, S.; Peters, W. Stereotactic localization and guidance using a machine vision technique. Stereotact. Funct. Neurosurg. 1992, 58, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Shams, S.; Butt, R.M.; Sarwar, A. Stereotactic biopsy of brain tumours. J.-Pak. Med. Assoc. 1996, 46, 176–177. [Google Scholar]
- Pennlund, A.; Jakola, A.S.; Skoglund, T.; Ljungqvist, J. A single-centre study of frame-based stereotactic brain biopsies. Br. J. Neurosurg. 2021, 10, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Catapano, G.; Sgulo, F.G.; Seneca, V.; Iorio, G.; de Notaris, M.; di Nuzzo, G. Fluorescein-assisted stereotactic needle biopsy of brain tumors: A single-center experience and systematic review. Neurosurg. Rev. 2019, 42, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Plunkett, R.; Allison, R.; Grand, W. Stereotactic neurosurgical biopsy is an underutilized modality. Neurosurg. Rev. 1999, 22, 117–120. [Google Scholar] [CrossRef]
- Kepes, J.J. Pitfalls and problems in the histopathologic evaluation of stereotactic needle biopsy specimens. Neurosurg. Clin. N. Am. 1994, 5, 19–33. [Google Scholar] [CrossRef]
- Engh, J.A.; Lunsford, L.D.; Amin, D.V.; Ochalski, P.G.; Fernandez-Miranda, J.; Prevedello, D.M.; Kassam, A.B. Stereotactically guided endoscopic port surgery for intraventricular tumor and colloid cyst resection. Oper. Neurosurg. 2010, 67, ons198–ons205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenfield, J.P.; Cobb, W.S.; Tsouris, A.J.; Schwartz, T.H. Stereotactic minimally invasive tubular retractor system for deep brain lesions. Oper. Neurosurg. 2008, 63, ONS334–ONS340. [Google Scholar] [CrossRef]
- Harris, A.E.; Hadjipanayis, C.G.; Lunsford, L.D.; Lunsford, A.K.; Kassam, A.B. Microsurgical removal of intraventricular lesions using endoscopic visualization and stereotactic guidance. Oper. Neurosurg. 2005, 56, ONS-125–ONS-132. [Google Scholar] [CrossRef]
- Ogura, K.; Tachibana, E.; Aoshima, C.; Sumitomo, M. New microsurgical technique for intraparenchymal lesions of the brain: Transcylinder approach. Acta Neurochir. 2006, 148, 779–785. [Google Scholar] [CrossRef]
- Romano, A.; Chibbaro, S.; Makiese, O.; Marsella, M.; Mainini, P.; Benericetti, E. Endoscopic removal of a central neurocytoma from the posterior third ventricle. J. Clin. Neurosci. 2009, 16, 312–316. [Google Scholar] [CrossRef]
- Kassam, A.B.; Engh, J.A.; Mintz, A.H.; Prevedello, D.M. Completely endoscopic resection of intraparenchymal brain tumors. J. Neurosurg. 2009, 110, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Margetis, K.; Souweidane, M.M. Endoscopic treatment of intraventricular cystic tumors. World Neurosurg. 2013, 79, S19.e1–S19.e11. [Google Scholar] [CrossRef] [PubMed]
- Jo, K.-I.; Chung, S.B.; Jo, K.-W.; Kong, D.-S.; Seol, H.-J.; Shin, H.-J. Microsurgical resection of deep-seated lesions using transparent tubular retractor: Pediatric case series. Child’s Nerv. Syst. 2011, 27, 1989. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R. History of neuroendoscopy. Neurosurg. Clin. 2004, 15, 1–7. [Google Scholar] [CrossRef]
- Decq, P.; Schroeder, H.W.; Fritsch, M.; Cappabianca, P. A history of ventricular neuroendoscopy. World Neurosurg. 2013, 79, S14.e11–S14.e16. [Google Scholar] [CrossRef]
- D’Angelo, L.; Armocida, D.; Sampirisi, L.; Paglia, F.; Berra, L.V.; Santoro, A. Role of endoscopic surgical biopsy in diagnoses of intraventricular/periventricular tumors: Review of literature including a monocentric case series. Acta Neurol. Belg. 2020, 120, 517–530. [Google Scholar] [CrossRef]
- Proctor, M.R.; Meara, J.G. A review of the management of single-suture craniosynostosis, past, present, and future: JNSPG 75th Anniversary Invited Review Article. J. Neurosurg. Pediatr. 2019, 24, 622–631. [Google Scholar] [CrossRef] [Green Version]
- Fries, G.; Perneczky, A. Intracranial endoscopy. Adv. Tech. Stand. Neurosurg. 1999, 6, 21–60. [Google Scholar]
- Perneczky, A.; Fries, G. Endoscope-assisted brain surgery: Part 1—Evolution, basic concept, and current technique. Neurosurgery 1998, 42, 219–224. [Google Scholar] [CrossRef]
- Brockmeyer, D. Techniques of endoscopic third ventriculostomy. Neurosurg. Clin. 2004, 15, 51–59. [Google Scholar] [CrossRef]
- Yokoh, A.; Sugita, K.; Kobayashi, S. Intermittent versus continuous brain retraction: An experimental study. J. Neurosurg. 1983, 58, 918–923. [Google Scholar] [CrossRef]
- Teo, C. Endoscopic-assisted tumor and neurovascular procedures. Clin. Neurosurg. 2000, 46, 515–525. [Google Scholar] [PubMed]
- Wilson, D.A.; Duong, H.; Teo, C.; Kelly, D.F. The supraorbital endoscopic approach for tumors. World Neurosurg. 2014, 82, e243–e256. [Google Scholar] [CrossRef] [PubMed]
- Stachura, K.; Grzywna, E.; Krzyżewski, R.M.; Kwinta, B.M.; Adamek, D.; Moskała, M.M. Endoscopic biopsy of intra-and paraventricular brain tumors. Videosurg. Other Miniinvasive Tech. 2019, 14, 107–113. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gradišnik, L.; Bošnjak, R.; Bunc, G.; Ravnik, J.; Maver, T.; Velnar, T. Neurosurgical Approaches to Brain Tissue Harvesting for the Establishment of Cell Cultures in Neural Experimental Cell Models. Materials 2021, 14, 6857. https://doi.org/10.3390/ma14226857
Gradišnik L, Bošnjak R, Bunc G, Ravnik J, Maver T, Velnar T. Neurosurgical Approaches to Brain Tissue Harvesting for the Establishment of Cell Cultures in Neural Experimental Cell Models. Materials. 2021; 14(22):6857. https://doi.org/10.3390/ma14226857
Chicago/Turabian StyleGradišnik, Lidija, Roman Bošnjak, Gorazd Bunc, Janez Ravnik, Tina Maver, and Tomaž Velnar. 2021. "Neurosurgical Approaches to Brain Tissue Harvesting for the Establishment of Cell Cultures in Neural Experimental Cell Models" Materials 14, no. 22: 6857. https://doi.org/10.3390/ma14226857
APA StyleGradišnik, L., Bošnjak, R., Bunc, G., Ravnik, J., Maver, T., & Velnar, T. (2021). Neurosurgical Approaches to Brain Tissue Harvesting for the Establishment of Cell Cultures in Neural Experimental Cell Models. Materials, 14(22), 6857. https://doi.org/10.3390/ma14226857