
Pathogenicity of GI-23 Avian Infectious Bronchitis Virus Strain Isolated in Brazil
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Disease
2.2. Viral Screening by Real-Time RT-PCR Assay
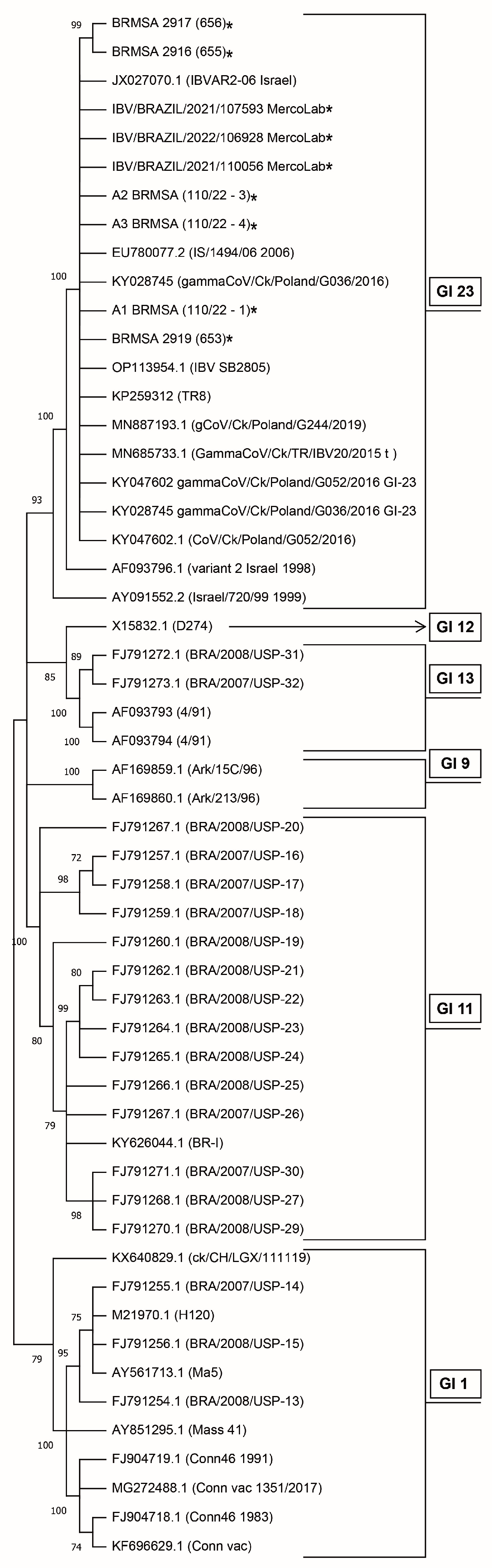
2.3. DNA Sequencing and Phylogenetic Analysis
2.4. Virus Isolation and Titration in Specific Pathogen-Free Chicken Embryonated Eggs (SPF-CEE)
2.5. Pathogenicity In Vivo Assays
2.5.1. Experimental Design
2.5.2. Ciliary Activity
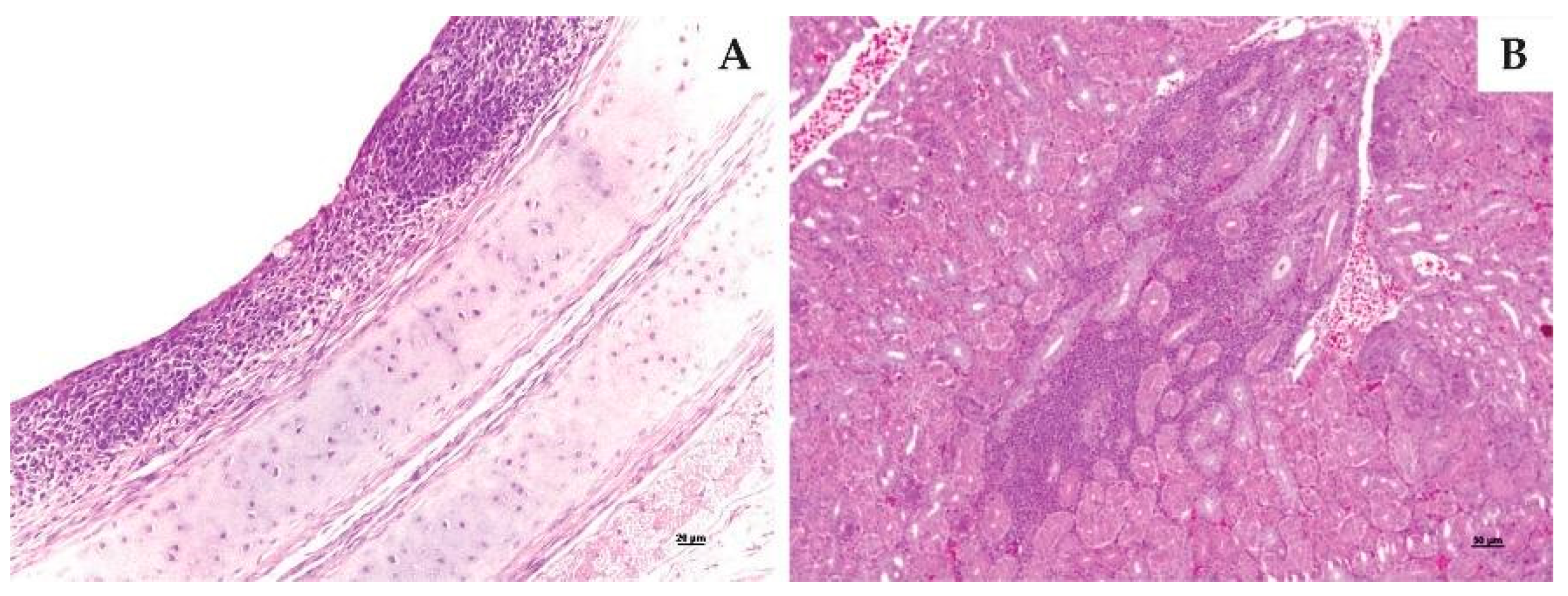
2.5.3. Histopathological Examination
2.6. Ethics Statement
3. Results
3.1. IBV Screening by RT-qPCR and Sequencing of GI-23 Lineage
3.2. Virus Isolation in SPF CEE
3.3. Pathogenicity In Vivo Assays
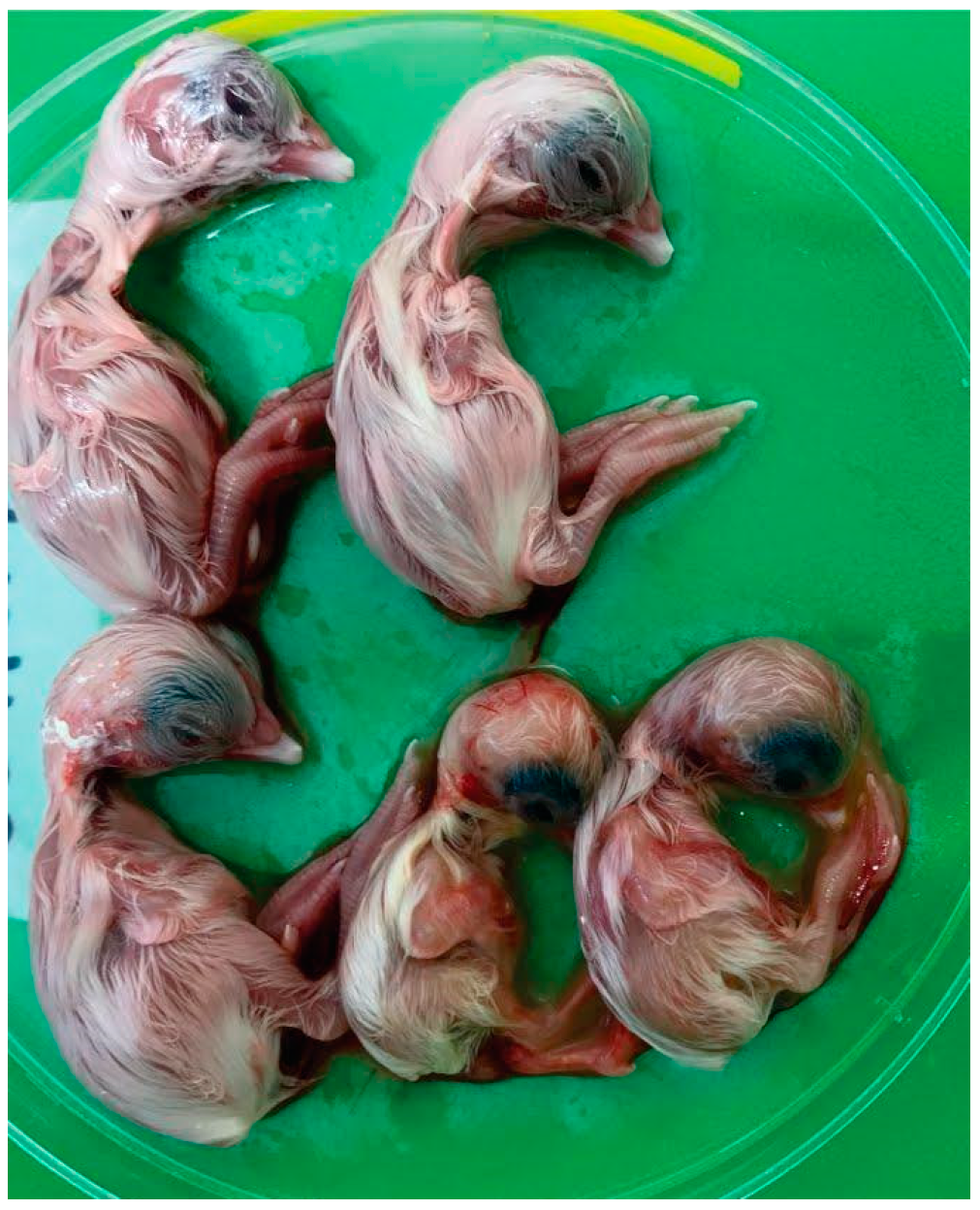
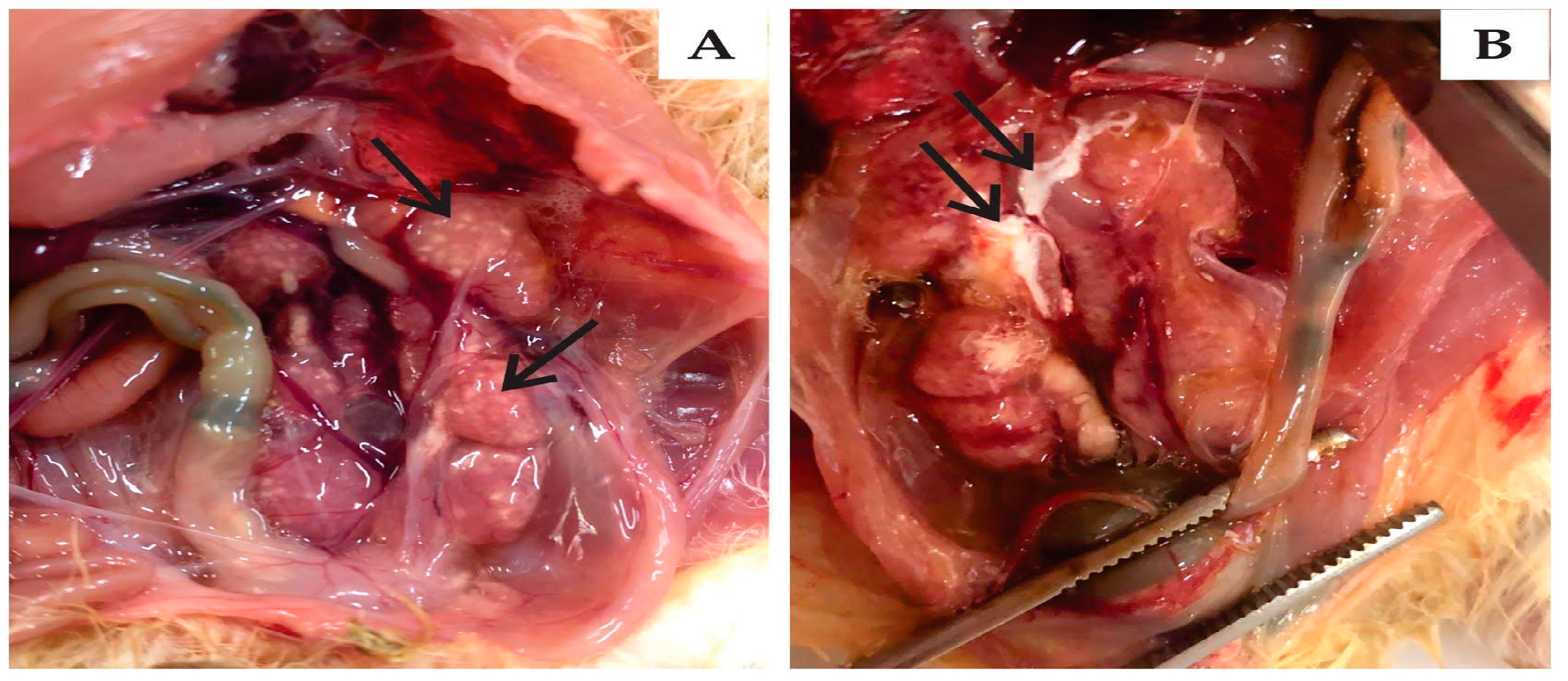
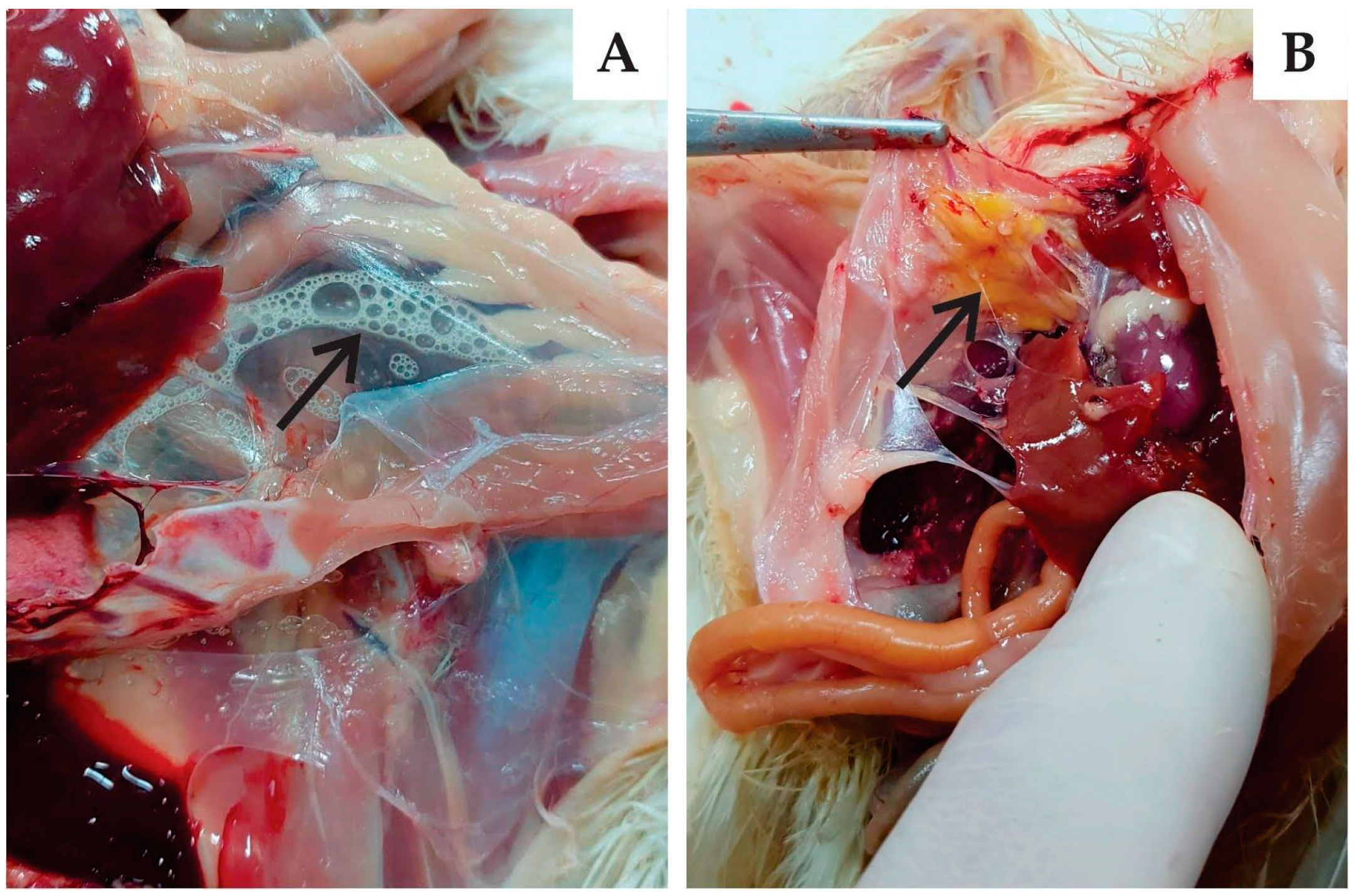
3.3.1. Clinical Signs and Gross Pathology
3.3.2. Ciliary Activity
3.3.3. Histopathological Examination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jackwood, M.W. Review of infectious bronchitis virus around the world. Avian Dis. 2012, 56, 634–641. [Google Scholar] [CrossRef]
- World Organisation for Animal Health—WOAH. High Pathogenicity Avian Influenza (HPAI)—Situation Report. Available online: https://www.woah.org/app/uploads/2023/03/hpai-situation-report-20230311.pdf (accessed on 25 March 2023).
- World Organisation for Animal Health—WOAH. Animal Disease Events: Country: Brazil. Disease: Newcastle Disease Virus (NCDV). Available online: https://wahis.woah.org/#/event-management (accessed on 25 March 2023).
- ICTV. Virus Taxonomy: Classification and Nomenclatute of Virusers: Ninth Report of the International Committee on Taxonomy of Viruses. 2018. Available online: https://talk.ictvonline.org//taxonomy/p/taxonomy-history?taxnode_id= (accessed on 25 March 2023).
- De Wit, J.J.S.; Cook, J.A. Spotlight on avian coronaviruses. Avian Pathol. 2020, 49, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W.; de Wit, S. Infectious Bronchitis-ch4. In Diseases of Poultry 14th Edition; Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Wit, S., Grimes, T., Johnson, D., Kromm, M., et al., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2020; pp. 167–188. [Google Scholar]
- Naqi, S.A.; Cowen, B.S.; Hattel, A.L.; Wilson, R.A. Detection of viral antigen following exposure of one-day-old chickens to the Holland 52 strain of infectious bronchitis virus. Avian Pathol. 1991, 20, 663–673. [Google Scholar] [CrossRef]
- Valastro, V.; Holmes, E.C.; Britton, P.; Fusaro, A.; Jackwood, M.W.; Cattoli, G.; Monne, I. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect. Genet. Evol. 2016, 39, 349–364. [Google Scholar] [CrossRef]
- Meir, R.; Rosenblut, E.; Perl, S.; Kass, N.; Ayali, G.; Hemsani, E.; Perk, S. Identification of a novel nephropathogenic infectious bronchitis virus in Israel. Avian Dis. 2004, 48, 635–641. [Google Scholar] [CrossRef]
- Kahya, S.; Coven, F.; Temelli, S.; Eyigor, A.; Carli, K.T. Presence of IS/1494/06 genotype-related infectious bronchitis virus in breeder and broiler flocks in breeder and broiler flocks in Turkey. Turk. Vet. J. 2013, 60, 27–31. [Google Scholar] [CrossRef]
- Selim, K.; Arafa, A.S.; Hussein, H.A.W.; El-Sanousi, A.A. Molecular characterisation of infectious bronchitis viruses isolated from broiler and layer chicken farms in Egypt during 2012. Int. J. Vet. Sci. Med. 2013, 1, 102–108. [Google Scholar] [CrossRef]
- Awad, F.; Baylis, M.; Ganapathy, K. Detection of variant infectious bronchitis viruses in boiler flocks in Libya. Int. J. Vet. Sci. Med. 2014, 2, 78–82. [Google Scholar] [CrossRef]
- Hussein, H.A.; Emara, M.M.; Rohaim, M.A.; Ganapathy, K.; Arafa, A.M. Sequence analysis of infectious bronchitis virus IS/1494 like strain isolated from broiler chicken co-infected with Newcastle disease virus in Egypt during 2012. Int. J. Poult. Sci. 2014, 13, 530–536. [Google Scholar] [CrossRef]
- Seger, W.; Langeroudi, A.G.; Karimi, V.; Madadgar, O.; Marandi, M.V.; Hashemzadeh, M. Prevalence of avian infectious bronchitis virus in broiler chicken farms in south of Iraq, 2014–2015. In Veterinary Research Forum; Faculty of Veterinary Medicine, Urmia University: Urmia, Iran, 2016; Volume 7, p. 317. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5251354/ (accessed on 20 January 2022).
- Yilmaz, H.; Altan, E.; Cizmecigil, U.Y.; Gurel, A.; Ozturk, G.Y.; Bamac, O.E.; Aydin, O.; Britton, P.; Monne, I.; Cetinkaya, B.; et al. Phylogeny and S1 gene variation of infectious bronchitis virus detected in broilers and layers in Turkey. Avian Dis. 2016, 60, 596–602. [Google Scholar] [CrossRef]
- Lisowska, A.; Sajewicz-Krukowska, J.; Fusaro, A.; Pikula, A.; Domanska-Blicharz, K. First characterization of a Middle-East GI-23 lineage (Var2-like) of infectious bronchitis virus in Europe. Virus Res. 2017, 242, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Müstak, I.B.; Müstak, H.K.; Bilgen, N. S1 gene based phylogeny of Israel variant-2 infectious bronchitis virus isolated in Turkey in a five year period. Pol. J. Vet. Sci. 2022, 25, 45–50. [Google Scholar]
- Fischer, S.; Klosterhalfen, D.; Wilms-Schulze, F.K.; Casteel, M. Research Note: First evidence of infectious bronchitis virus Middle-East GI-23 lineage (Var2-like) in Germany. Poult. Sci. 2020, 99, 797–800. [Google Scholar] [CrossRef] [PubMed]
- Houta, M.H.; Hassan, K.E.; El-Sawah, A.A.; Elkady, M.F.; Kilany, W.H.; Ali, A.; Abdel-Moneim, A.S. The emergence, evolution and spread of infectious bronchitis virus genotype GI-23. Arch. Virol. 2021, 166, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Chacón, J.L.; Rodrigues, J.N.; Júnior, M.S.A.; Peloso, C.; Pedroso, A.C.; Ferreira, A.J.P. Epidemiological survey and molecular characterization of avian infectious bronchitis virus in Brazil between 2003 and 2009. Avian Pathol. 2011, 40, 153–162. [Google Scholar] [CrossRef]
- Fernando, F.S.; Kasmanas, T.C.; Lopes, P.D.; Montassier, M.F.D.; Mores, M.A.Z.; Casagrande Mariguela, V.; Pavani, C.; Moreira Dos Santos, R.; Assayag, M.S., Jr.; Montassier, H.J. Assessment of molecular and genetic evolution, antigenicity and virulence properties during the persistence of the infectious bronchitis virus in broiler breeders. J. Gen. Virol. 2017, 98, 2470–2481. [Google Scholar] [CrossRef]
- Fraga, A.P.; Balestrin, E.; Ikuta, N.; Fonseca, A.S.K.; Spilki, F.R.; Canal, C.W.; Lunge, V.R. Emergence of a new genotype of avian infectious bronchitis virus in Brazil. Avian Dis. 2013, 57, 225–232. [Google Scholar] [CrossRef]
- Fraga, A.P.; de Gräf, T.; Pereira, C.S.; Ikuta, N.; Fonseca, A.S.K.; Lunge, V.R. Phylodynamic analysis and molecular diversity of the avian infectious bronchitis virus of chickens in Brazil. Infect. Genet. Evol. 2018, 59, 155–162. [Google Scholar] [CrossRef]
- Marandino, A.; Pereda, A.; Tomás, G.; Hernández, M.; Iraola, G.; Craig, M.I.; Hernaández, D.; Banda, A.; Villegas, P.; Panzera, Y.; et al. Phylodynamic analysis of avian infectious bronchitis virus in South America. J. Gen. Virol. 2015, 96 Pt 6, 1340. [Google Scholar] [CrossRef]
- Domanska-Blicharz, K.; Jacukowicz, A.; Lisowska, A.; Wyrostek, K.; Minta, Z. Detection and molecular characterization of infectious bronchitis-like viruses in wild bird populations. Avian Pathol. 2014, 43, 406–413. [Google Scholar] [CrossRef]
- Ikuta, N.; Fonseca, A.S.K.; Fernando, F.S.; Filho, T.F.; Martins, N.R.S.; Lunge, V.R. Emergence and molecular characterization of the avian infectious bronchitis virus GI-23 in commercial broiler farms from South America. Anim. Health Res. Rev. 2022, 69, 3167–3172. [Google Scholar] [CrossRef] [PubMed]
- Callison, S.A.; Hilt, D.A.; Boynton, T.O.; Sample, B.F.; Robison, R.; Swayne, D.E.; Jackwood, M.W. Development and evaluation of a real-time Taqman RT-PCR assay for the detection of infectious bronchitis virus from infected chickens. J. Virol. Methods 2006, 138, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Fraga, A.P.; Ikuta, N.; Fonseca, A.S.K.; Spilki, F.R.; Balestrin, E.; Rodrigues, C.D.; Canal, W.C.; Lunge, V.R. A real-time reverse-530 transcription polymerase chain reaction for differentiation of Massachusetts vaccine and Brazilian field genotypes of avian infectious 531 bronchitis virus. Avian Dis. 2016, 60, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.I.; Carlsbad, C. BioEdit: An Important Software for Molecular Biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Gelb, J., Jr. A Laboratory Manual for the Isolation and Identification of Avian Pathogens, 3rd ed.; Purchase, H.G., Arp, L.H., Domermuth, C.H., Pearson, J.E., Eds.; Infectious Bronchitis; American Association of Avian Pathologists, University of Pennsylvania: Kennett Square, PA, USA, 1998; Chapter 29; pp. 124–127. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent end points. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Ignjatovic, J.; Gould, G.; Sapats, S. Isolation of a variant infectious bronchitis virus in Australia that further illustrates diversity among emerging strains. Arch. Virol. 2006, 151, 1567–1585. [Google Scholar] [CrossRef]
- Cavanagh, D.; Elus, M.M.; Cook, J.K.A. Relationship between sequence variation in the S1 spike protein of infectious bronchitis virus and the extent of cross-protection in vivo. Avian Pathol. 1997, 26, 63–74. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Zanaty, A.; Naguib, M.M.; El-Husseiny, M.H.; Mady, W.; Hagag, N.; Arafa, A.S. The sequence of the full spike S1 glycoprotein of infectious bronchitis virus circulating in Egypt reveals evidence of intra-genotypic recombination. Arch. Virol. 2016, 161, 3583–3587. [Google Scholar] [CrossRef]
- Lisowska, A.; Pikuła, A.; Opolska, J.; Jasik, A.; Kycko, A.; Domańska-Blicharz, K. Virulence Properties of GI-23 Infectious Bronchitis Virus Isolated in Poland and Efficacy of Different Vaccination strategies. Pathogens 2021, 10, 522. [Google Scholar] [CrossRef] [PubMed]
- Awad, F.; Chhabra, R.; Forrester, A.; Chantrey, J.; Baylis, M.; Lemiere, S.; Hussein, H.A.; Ganapathy, K. Experimental infection of IS/885/00-like infectious bronchitis virus in specific pathogen free and commercial broiler chicks. Res. Vet. Sci. 2016, 105, 15–22. [Google Scholar] [CrossRef] [PubMed]
- OIE Terrestrial Manual. Avian infectious bronchitis (BI). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 6th ed.; Office International des Epizooties: Paris, France, 2008; Chapter 2.3.2; pp. 443–455. [Google Scholar]
- Butcher, G.D.; Winterfield, B.; Shapiroc, D.P. Pathogenesis of H13 Nephropathogenic Infectious Bronchitis Virus. Avian Dis. 1990, 34, 916–921. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus/Strain * | Age (Days) | Organ Isolated | Symptom | Year of Isolation |
|---|---|---|---|---|
| IBV-BRMSA2916 (655) | 23 | Cecal tonsil | Respiratory | 2021 |
| IBV-BRMSA2917 (656) | 41 | Trachea | Respiratory | 2021 |
| IBV-BRMSA2919 (653) | 23 | Cecal tonsil | Respiratory | 2021 |
| IBV-BRMSA3090 (110-3) | 27 | Kidney | Respiratory | 2022 |
| IBV-BRMSA3091 (110-4) | 27 | Kidney | Respiratory | 2022 |
| IBV-BRMSA3092 (110-1) | 15 | Lung | Respiratory | 2022 |
| Group | Tissues | Score and Percentage/Group * | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 6 dpi | 13 dpi | ||||||||||
| 0 | 1 | 2 | 3 | % | 0 | 1 | 2 | 3 | % | ||
| Negative Control (NC) | CA | 4 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 |
| T | 4 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | |
| K | 4 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | |
| BRMSA2916 (655) | CA | 0 | 0 | 0 | 6 | 100 | 0 | 2 | 2 | 2 | 67 |
| T | 0 | 2 | 4 | 0 | 67 | 0 | 2 | 3 | 0 | 60 | |
| K | 0 | 1 | 2 | 3 | 83 | 1 | 3 | 2 | 0 | 33 | |
| BRMSA2917 (656) | CA | 0 | 0 | 0 | 6 | 100 | 0 | 2 | 2 | 2 | 67 |
| T | 0 | 1 | 3 | 2 | 83 | 0 | 2 | 2 | 1 | 50 | |
| K | 3 | 0 | 2 | 1 | 50 | 2 | 2 | 1 | 1 | 33 | |
| BRMSA2919 (653) | CA | 0 | 0 | 0 | 6 | 100 | 0 | 0 | 3 | 3 | 100 |
| T | 0 | 0 | 2 | 4 | 100 | 0 | 2 | 3 | 1 | 67 | |
| K | 3 | 2 | 0 | 1 | 17 | 4 | 2 | 1 | 0 | 17 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trevisol, I.M.; Caron, L.; Mores, M.A.Z.; Voss-Rech, D.; da Silva Zani, G.; Back, A.; Marchesi, J.A.P.; Esteves, P.A. Pathogenicity of GI-23 Avian Infectious Bronchitis Virus Strain Isolated in Brazil. Viruses 2023, 15, 1200. https://doi.org/10.3390/v15051200
Trevisol IM, Caron L, Mores MAZ, Voss-Rech D, da Silva Zani G, Back A, Marchesi JAP, Esteves PA. Pathogenicity of GI-23 Avian Infectious Bronchitis Virus Strain Isolated in Brazil. Viruses. 2023; 15(5):1200. https://doi.org/10.3390/v15051200
Chicago/Turabian StyleTrevisol, Iara Maria, Luizinho Caron, Marcos Antônio Zanella Mores, Daiane Voss-Rech, Gabriel da Silva Zani, Alberto Back, Jorge Augusto Petroli Marchesi, and Paulo Augusto Esteves. 2023. "Pathogenicity of GI-23 Avian Infectious Bronchitis Virus Strain Isolated in Brazil" Viruses 15, no. 5: 1200. https://doi.org/10.3390/v15051200
APA StyleTrevisol, I. M., Caron, L., Mores, M. A. Z., Voss-Rech, D., da Silva Zani, G., Back, A., Marchesi, J. A. P., & Esteves, P. A. (2023). Pathogenicity of GI-23 Avian Infectious Bronchitis Virus Strain Isolated in Brazil. Viruses, 15(5), 1200. https://doi.org/10.3390/v15051200