
Association between Arachidonic Acid and the Risk of Schizophrenia: A Cross-National Study and Mendelian Randomization Analysis

 ,
,
Abstract
:1. Introduction
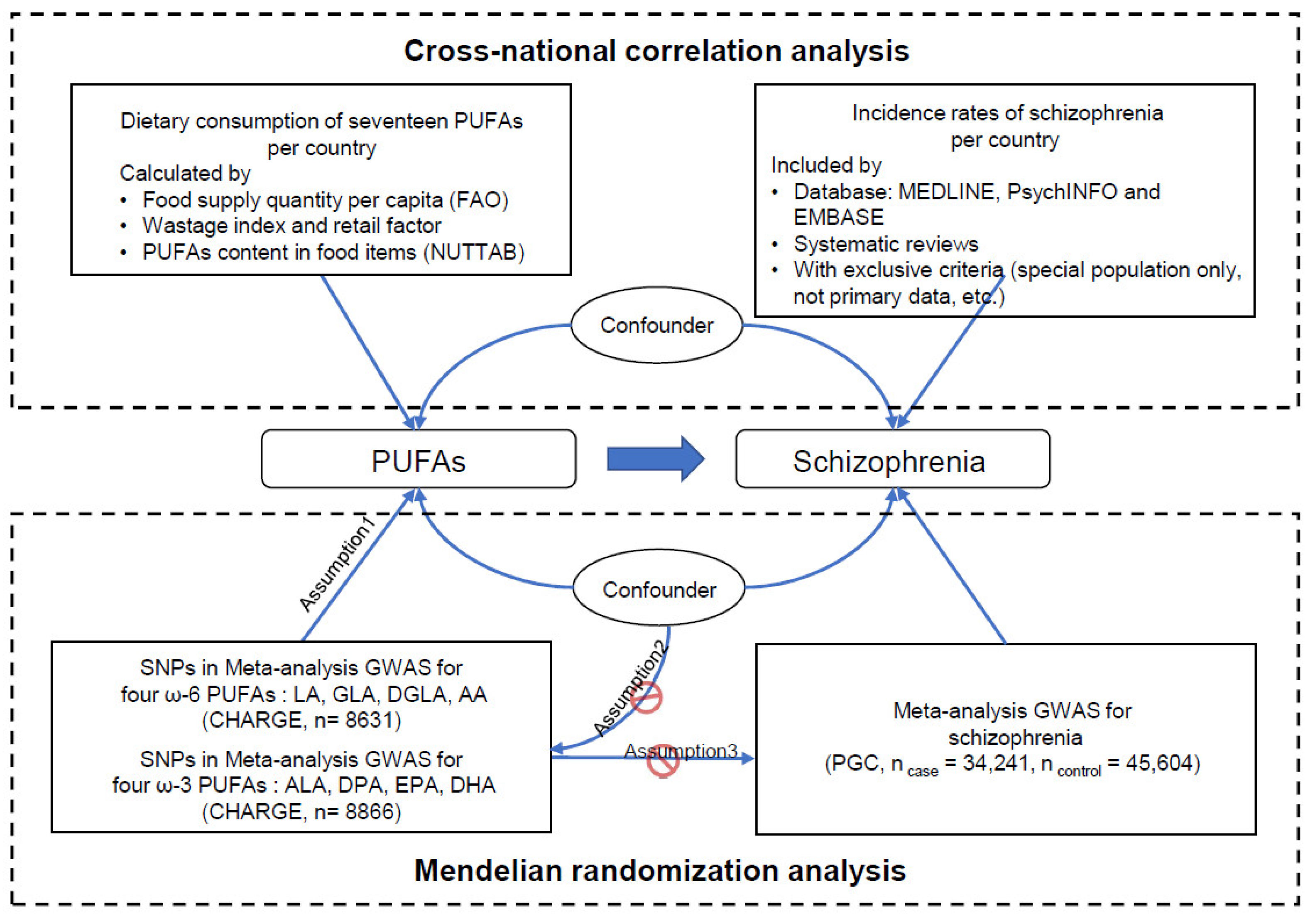
2. Materials and Methods
2.1. Cross-National Correlation between PUFA Consumption and Schizophrenia Incidence Rates
2.1.1. Data Acquisition on the Incidence Rates of Schizophrenia
2.1.2. Data Acquisition on the Dietary Consumption of PUFAs
2.1.3. Correlation Adjustment
2.1.4. Statistical Analysis
2.2. Mendelian Randomization Analysis for PUFAs and Schizophrenia
2.2.1. PUFA Exposure Data Acquisition
2.2.2. Schizophrenia Outcome Data Acquisition
2.2.3. Univariable MR Analysis
2.2.4. Multivariable MR Analysis
2.2.5. Pleiotropic Associations
3. Results
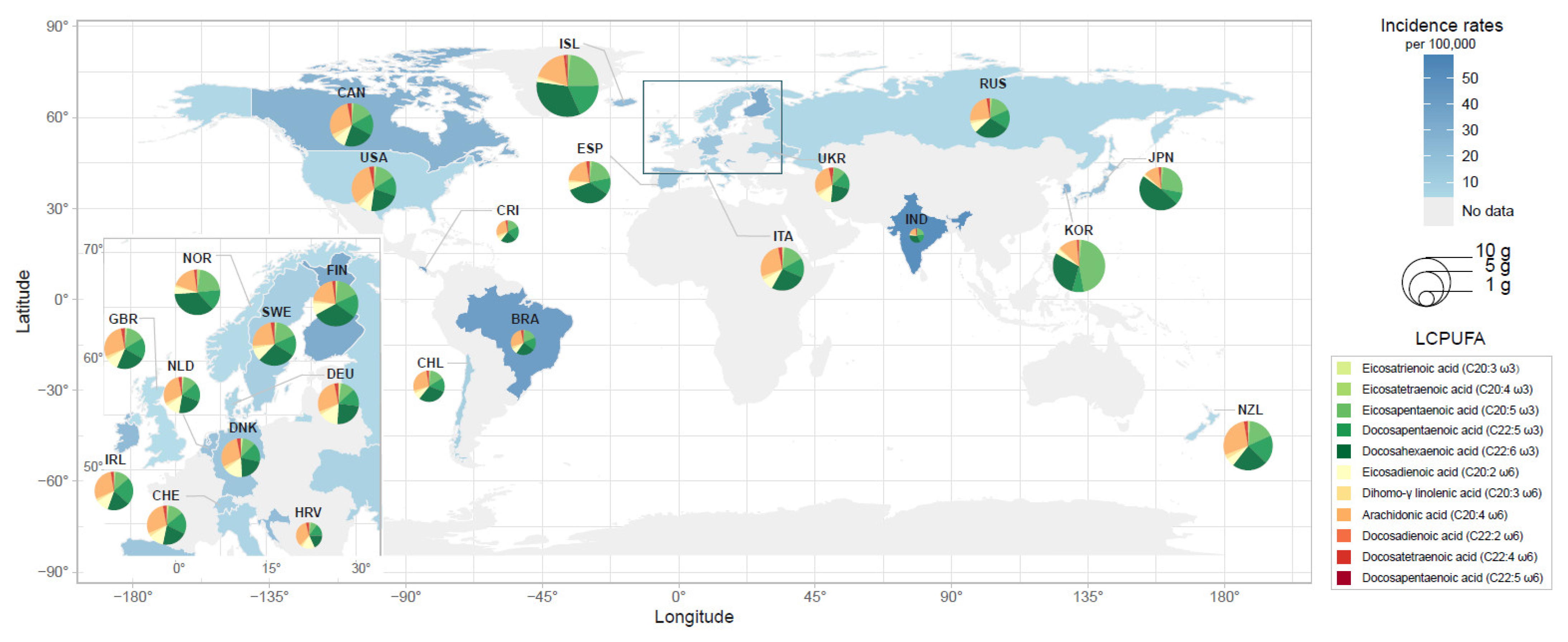
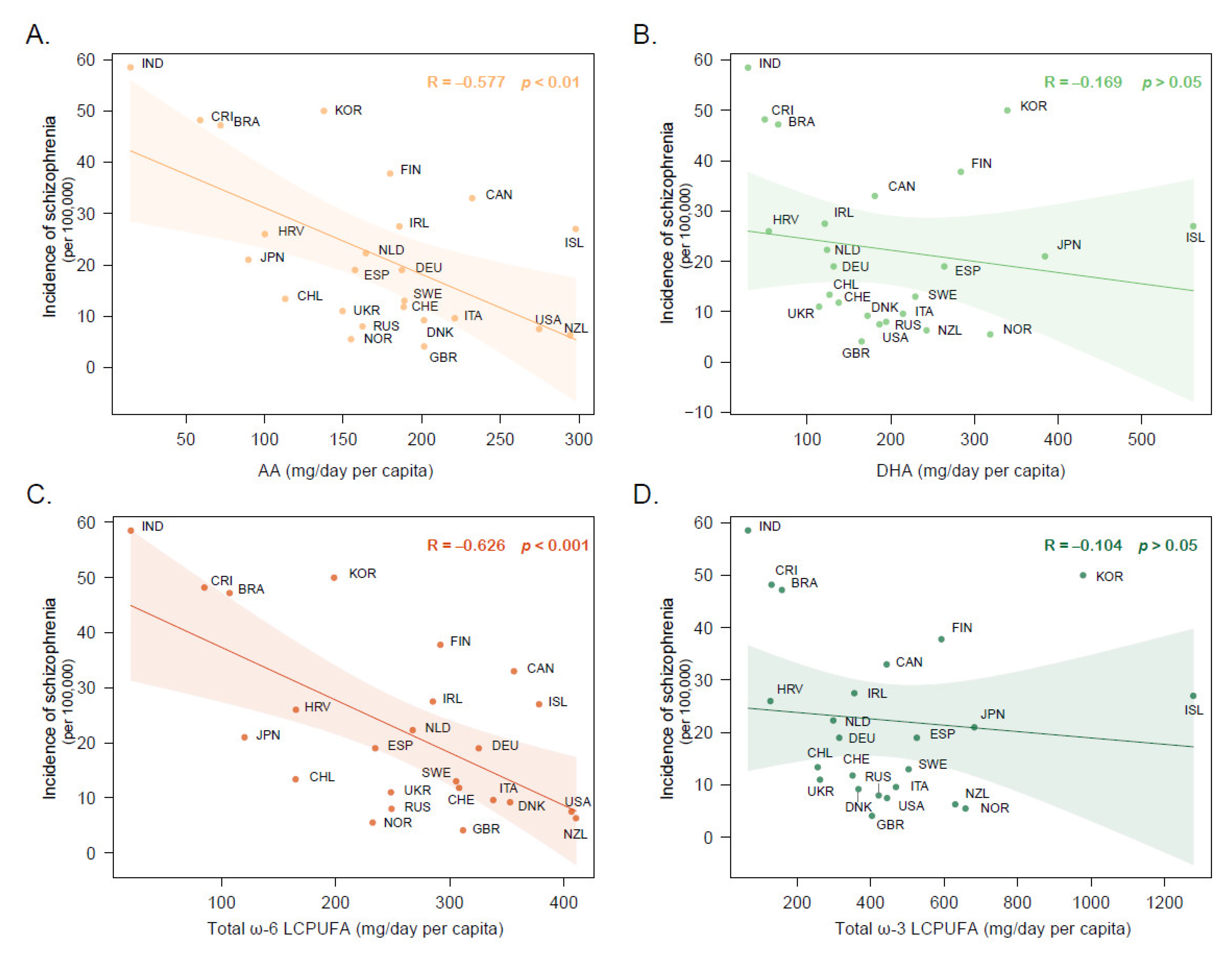
3.1. Cross-National Correlation between Dietary PUFA Intake and Schizophrenia Incidence Rates
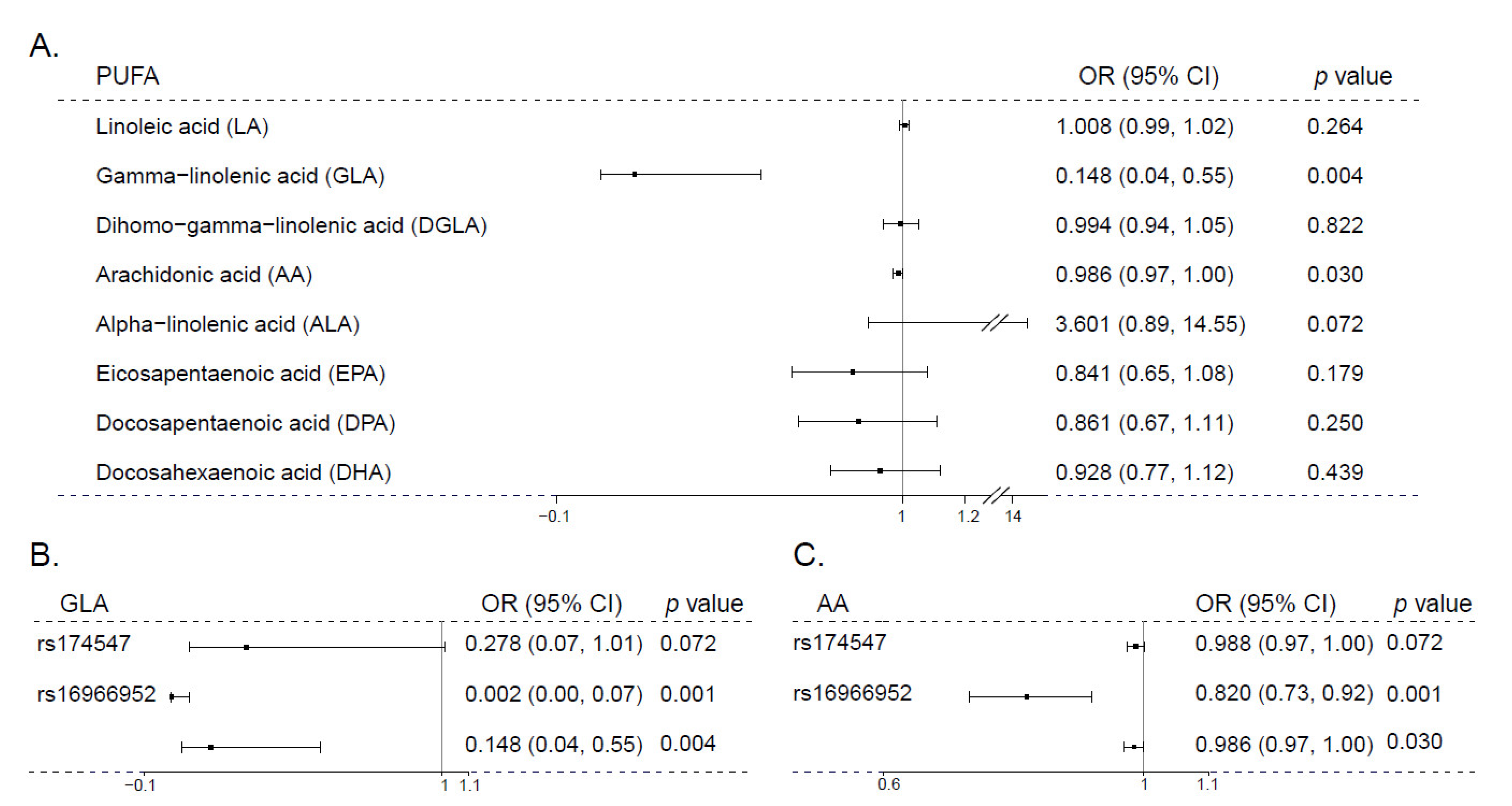
3.2. Mendelian Randomization Analysis of PUFA and Schizophrenia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mueser, K.T.; McGurk, S.R. Schizophrenia. Lancet 2004, 363, 2063–2072. [Google Scholar] [CrossRef]
- McGrath, J.; Saha, S.; Welham, J.; El Saadi, O.; MacCauley, C.; Chant, D. A systematic review of the incidence of schizophrenia: The distribution of rates and the influence of sex, urbanicity, migrant status and methodology. BMC Med. 2004, 2, 13. [Google Scholar] [CrossRef] [Green Version]
- Lewis, L. Mood disorders: Diagnosis, treatment, and support from a patient perspective. Psychopharmacol. Bull. 2001, 35, 186–196. [Google Scholar]
- Susser, E.; Neugebauer, R.; Hoek, H.W.; Brown, A.S.; Lin, S.; Labovitz, D.; Gorman, J.M. Schizophrenia after prenatal famine. Further evidence. Arch. Gen. Psychiatry 1996, 53, 25–31. [Google Scholar] [CrossRef] [PubMed]
- St Clair, D.; Xu, M.; Wang, P.; Yu, Y.; Fang, Y.; Zhang, F.; Zheng, X.; Gu, N.; Feng, G.; Sham, P.; et al. Rates of adult schizophrenia following prenatal exposure to the Chinese famine of 1959–1961. JAMA 2005, 294, 557–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muskiet, F.A.J. Pathophysiology and Evolutionary Aspects of Dietary Fats and Long-Chain Polyunsaturated Fatty Acids across the Life Cycle. In Fat Detection: Taste, Texture, and Post Ingestive Effects; Frontiers in Neuroscience; Montmayeur, J.P., le Coutre, J., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2010. [Google Scholar]
- Janssen, C.I.; Kiliaan, A.J. Long-chain polyunsaturated fatty acids (LCPUFA) from genesis to senescence: The influence of LCPUFA on neural development, aging, and neurodegeneration. Prog. Lipid. Res. 2014, 53, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lepping, R.J.; Honea, R.A.; Martin, L.E.; Liao, K.; Choi, I.Y.; Lee, P.; Papa, V.B.; Brooks, W.M.; Shaddy, D.J.; Carlson, S.E.; et al. Long-chain polyunsaturated fatty acid supplementation in the first year of life affects brain function, structure, and metabolism at age nine years. Dev. Psychobiol. 2019, 61, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birch, E.E.; Garfield, S.; Castaneda, Y.; Hughbanks-Wheaton, D.; Uauy, R.; Hoffman, D. Visual acuity and cognitive outcomes at 4 years of age in a double-blind, randomized trial of long-chain polyunsaturated fatty acid-supplemented infant formula. Early Hum. Dev. 2007, 83, 279–284. [Google Scholar] [CrossRef]
- Willatts, P.; Forsyth, S.; Agostoni, C.; Casaer, P.; Riva, E.; Boehm, G. Effects of long-chain PUFA supplementation in infant formula on cognitive function in later childhood. Am. J. Clin. Nutr. 2013, 98, 536S–542S. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.D.; Jones, H.J.; Heron, J.; Hibbeln, J.; Sullivan, S.; Zammit, S. Omega-3 and Omega-6 fatty acids and risk of psychotic outcomes in the ALSPAC birth cohort. Schizophr. Res. 2020, 224, 108–115. [Google Scholar] [CrossRef]
- Hoen, W.P.; Lijmer, J.G.; Duran, M.; Wanders, R.J.; van Beveren, N.J.; de Haan, L. Red blood cell polyunsaturated fatty acids measured in red blood cells and schizophrenia: A meta-analysis. Psychiatry Res. 2013, 207, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Van der Kemp, W.J.; Klomp, D.W.; Kahn, R.S.; Luijten, P.R.; Hulshoff Pol, H.E. A meta-analysis of the polyunsaturated fatty acid composition of erythrocyte membranes in schizophrenia. Schizophr. Res. 2012, 141, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Alqarni, A.; Mitchell, T.W.; McGorry, P.D.; Nelson, B.; Markulev, C.; Yuen, H.P.; Schafer, M.R.; Berger, M.; Mossaheb, N.; Schlogelhofer, M.; et al. Comparison of erythrocyte omega-3 index, fatty acids and molecular phospholipid species in people at ultra-high risk of developing psychosis and healthy people. Schizophr. Res. 2020, 226, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Charlson, F.J.; Ferrari, A.J.; Santomauro, D.F.; Diminic, S.; Stockings, E.; Scott, J.G.; McGrath, J.J.; Whiteford, H.A. Global Epidemiology and Burden of Schizophrenia: Findings From the Global Burden of Disease Study 2016. Schizophr. Bull. 2018, 44, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Hogerzeil, S.J.; van Hemert, A.M. Design choices when estimating the treated incidence of schizophrenia. Psychol. Med. 2019, 49, 2635–2636. [Google Scholar] [CrossRef] [Green Version]
- Schmidhuber, J.; Sur, P.; Fay, K.; Huntley, B.; Salama, J.; Lee, A.; Cornaby, L.; Horino, M.; Murray, C.; Afshin, A. The Global Nutrient Database: Availability of macronutrients and micronutrients in 195 countries from 1980 to 2013. Lancet Planet Health 2018, 2, e353–e368. [Google Scholar] [CrossRef] [Green Version]
- Forsyth, S.; Gautier, S.; Salem, N., Jr. Global Estimates of Dietary Intake of Docosahexaenoic Acid and Arachidonic Acid in Developing and Developed Countries. Ann. Nutr. Metab. 2016, 68, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Lassek, W.D.; Gaulin, S.J. Linoleic and docosahexaenoic acids in human milk have opposite relationships with cognitive test performance in a sample of 28 countries. Prostaglandins Leukot. Essent. Fat. Acids 2014, 91, 195–201. [Google Scholar] [CrossRef]
- Basu, S.; McKee, M.; Galea, G.; Stuckler, D. Relationship of soft drink consumption to global overweight, obesity, and diabetes: A cross-national analysis of 75 countries. Am. J. Public Health 2013, 103, 2071–2077. [Google Scholar] [CrossRef]
- Fox, A.; Feng, W.; Asal, V. What is driving global obesity trends? Globalization or “modernization”? Glob. Health 2019, 15, 32. [Google Scholar] [CrossRef] [Green Version]
- Malik, V.S.; Willett, W.C.; Hu, F.B. Global obesity: Trends, risk factors and policy implications. Nat. Rev. Endocrinol. 2013, 9, 13–27. [Google Scholar] [CrossRef]
- Guan, W.; Steffen, B.T.; Lemaitre, R.N.; Wu, J.H.Y.; Tanaka, T.; Manichaikul, A.; Foy, M.; Rich, S.S.; Wang, L.; Nettleton, J.A.; et al. Genome-wide association study of plasma N6 polyunsaturated fatty acids within the cohorts for heart and aging research in genomic epidemiology consortium. Circ. Cardiovasc. Genet. 2014, 7, 321–331. [Google Scholar] [CrossRef] [Green Version]
- Lemaitre, R.N.; Tanaka, T.; Tang, W.; Manichaikul, A.; Foy, M.; Kabagambe, E.K.; Nettleton, J.A.; King, I.B.; Weng, L.C.; Bhattacharya, S.; et al. Genetic loci associated with plasma phospholipid n-3 fatty acids: A meta-analysis of genome-wide association studies from the CHARGE Consortium. PLoS Genet. 2011, 7, e1002193. [Google Scholar] [CrossRef] [Green Version]
- Ghoneim, D.H.; Zhu, J.; Zheng, W.; Long, J.; Murff, H.J.; Ye, F.; Setiawan, V.W.; Wilkens, L.R.; Khankari, N.K.; Haycock, P.; et al. Mendelian Randomization Analysis of n-6 Polyunsaturated Fatty Acid Levels and Pancreatic Cancer Risk. Cancer Epidemiol. Biomark. Prev. 2020, 29, 2735–2739. [Google Scholar] [CrossRef]
- Yuan, S.; Larsson, S.C. Association of genetic variants related to plasma fatty acids with type 2 diabetes mellitus and glycaemic traits: A Mendelian randomisation study. Diabetologia 2020, 63, 116–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khankari, N.K.; Banbury, B.L.; Borges, M.C.; Haycock, P.; Albanes, D.; Arndt, V.; Berndt, S.I.; Bezieau, S.; Brenner, H.; Campbell, P.T.; et al. Mendelian Randomization of Circulating Polyunsaturated Fatty Acids and Colorectal Cancer Risk. Cancer Epidemiol. Biomark. Prev. 2020, 29, 860–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomata, Y.; Larsson, S.C.; Hagg, S. Polyunsaturated fatty acids and risk of Alzheimer’s disease: A Mendelian randomization study. Eur. J. Nutr. 2020, 59, 1763–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schizophrenia Working Group of the Psychiatric Genomics Consortium. Biological insights from 108 schizophrenia-associated genetic loci. Nature 2014, 511, 421–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, S.; Thompson, S.G.; Collaboration, C.C.G. Avoiding bias from weak instruments in Mendelian randomization studies. Int. J. Epidemiol. 2011, 40, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, B.S.; Steiner, J.; Bernstein, H.G.; Dodd, S.; Pasco, J.A.; Dean, O.M.; Nardin, P.; Goncalves, C.A.; Berk, M. C-reactive protein is increased in schizophrenia but is not altered by antipsychotics: Meta-analysis and implications. Mol. Psychiatry 2016, 21, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Dickson, H.; Hedges, E.P.; Ma, S.Y.; Cullen, A.E.; MacCabe, J.H.; Kempton, M.J.; Downs, J.; Laurens, K.R. Academic achievement and schizophrenia: A systematic meta-analysis. Psychol. Med. 2020, 50, 1949–1965. [Google Scholar] [CrossRef] [PubMed]
- Radua, J.; Ramella-Cravaro, V.; Ioannidis, J.P.A.; Reichenberg, A.; Phiphopthatsanee, N.; Amir, T.; Yenn Thoo, H.; Oliver, D.; Davies, C.; Morgan, C.; et al. What causes psychosis? An umbrella review of risk and protective factors. World Psychiatry 2018, 17, 49–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seviiri, M.; Law, M.H.; Ong, J.S.; Gharahkhani, P.; Nyholt, D.R.; Olsen, C.M.; Whiteman, D.C.; MacGregor, S. Polyunsaturated Fatty Acid Levels and the Risk of Keratinocyte Cancer: A Mendelian Randomization Analysis. Cancer Epidemiol. Biomark. Prev. 2021, 30, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Machiela, M.J.; Chanock, S.J. LDlink: A web-based application for exploring population-specific haplotype structure and linking correlated alleles of possible functional variants. Bioinformatics 2015, 31, 3555–3557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ligthart, S.; Vaez, A.; Vosa, U.; Stathopoulou, M.G.; de Vries, P.S.; Prins, B.P.; Van der Most, P.J.; Tanaka, T.; Naderi, E.; Rose, L.M.; et al. Genome Analyses of >200,000 Individuals Identify 58 Loci for Chronic Inflammation and Highlight Pathways that Link Inflammation and Complex Disorders. Am. J. Hum. Genet. 2018, 103, 691–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamat, M.A.; Blackshaw, J.A.; Young, R.; Surendran, P.; Burgess, S.; Danesh, J.; Butterworth, A.S.; Staley, J.R. PhenoScanner V2: An expanded tool for searching human genotype-phenotype associations. Bioinformatics 2019, 35, 4851–4853. [Google Scholar] [CrossRef] [Green Version]
- Messias, E.; Sampaio, J.J.; Messias, N.C.; Kirkpatrick, B. Epidemiology of schizophrenia in northeast Brazil. J. Nerv. Ment. Dis. 2000, 188, 118–120. [Google Scholar] [CrossRef]
- Whitehorn, D.; Richard, J.C.; Kopala, L.C. Hospitalization in the first year of treatment for schizophrenia. Can. J. Psychiatry 2004, 49, 635–638. [Google Scholar] [CrossRef] [Green Version]
- Larach, V.; Varela, L.F.; Vergara, G.; Rojas, I.; Hernández, V.; Casanueva, R.; Irarrázaval, M. Universal healthcare coverage for first episode of schizophrenia-spectrum disorders in Chile: Analysis of the administrative database. Eur. Arch. Psychiatry Clin. Neurosci. 2022, 272, 129–138. [Google Scholar] [CrossRef]
- Handal, N.; Dodds, J.H. Statistics for initial admission for schizophrenia in hospitals in Costa Rica. Rev. Panam Salud Publica 1997, 1, 426–434. [Google Scholar]
- Folnegović, Z.; Folnegović-Šmalc, V.; Kulčar, Ž. The incidence of schizophrenia in Croatia. Br. J. Psychiatry 1990, 156, 363–365. [Google Scholar] [CrossRef]
- Castagnini, A.; Foldager, L. Variations in incidence and age of onset of acute and transient psychotic disorders. Soc. Psychiatry Psychiatr. Epidemiol. 2013, 48, 1917–1922. [Google Scholar] [CrossRef] [PubMed]
- Salokangas, R.K.R.; Helminen, M.; Koivisto, A.-M.; Rantanen, H.; Oja, H.; Pirkola, S.; Wahlbeck, K.; Joukamaa, M. Incidence of hospitalised schizophrenia in Finland since 1980: Decreasing and increasing again. Soc. Psychiatry Psychiatr. Epidemiol. 2011, 46, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Löffler, W.; Häfner, H. Ecological pattern of first admitted schizophrenics in two German cities over 25 years. Soc. Sci. Med. 1999, 49, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Helgason, L. Psychiatric services and mental illness in Iceland. Incidence study (1966–1967) with 6–7 year follow-up. Acta Psychiatr. Scand. Suppl. 1977, 268, 1–140. [Google Scholar]
- Rajkumar, S.; Padmavathi, R.; Thara, R.; Menon, M.S. Incidence of schizophrenia in an urban community in madras. Indian J. Psychiatry 1993, 35, 18–21. [Google Scholar] [PubMed]
- Blake, B.; Halpenny, J.V.; ni Nuallain, M.; O’Brien, P.F.; O’Hare, A.; Walsh, D. The incidence of mental illness in Ireland—Patients contacting psychiatric services in three Irish Counties. Ir. J. Psychiatry 1984, 5, 23–29. [Google Scholar]
- Mulè, A.; Sideli, L.; Capuccio, V.; Fearon, P.; Ferraro, L.; Kirkbride, J.B.; La Cascia, C.; Sartorio, C.; Seminerio, F.; Tripoli, G.; et al. Low incidence of psychosis in Italy: Confirmation from the first epidemiological study in Sicily. Soc. Psychiatry Psychiatr. Epidemiol. 2017, 52, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Ohta, Y.; Nakane, Y.; Nishihara, J.; Takemoto, T. Ecological structure and incidence rates of schizophrenia in Nagasaki City. Acta Psychiatr. Scand. 1992, 86, 113–120. [Google Scholar] [CrossRef]
- Van Os, J.; Driessen, G.; Gunther, N.; Delespaul, P. Neighbourhood variation in incidence of schizophrenia. Evidence for person-environment interaction. Br. J. Psychiatry 2000, 176, 243–248. [Google Scholar] [CrossRef]
- Tapsell, R.; Hallett, C.; Mellsop, G. The rate of mental health service use in New Zealand as analysed by ethnicity. Australas Psychiatry 2018, 26, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Johannessen, J.O. Schizofreni-forekomst i et norsk fylke, Rogaland = The occurrence of schizophrenia in the Norwegian county of Rogaland. Nord. Psykiatr. Tidsskr. 1985, 39, 217–223. [Google Scholar] [CrossRef]
- Jung, Y.S.; Kim, Y.E.; Go, D.S.; Yoon, S.J. The prevalence, incidence, and admission rate of diagnosed schizophrenia spectrum disorders in Korea, 2008–2017: A nationwide population-based study using claims big data analysis. PLoS ONE 2021, 16, e0256221. [Google Scholar] [CrossRef] [PubMed]
- Kazakovtsev, B.A.; Sidoryuk, O.V.; Zrazhevskaya, I.A.; Ovsyannikov, S.A. Regional differences in the incidence of affective disorders. Zh. Nevrol. Psikhiatr. Im S S Korsakova 2019, 119, 10–16. [Google Scholar] [CrossRef]
- Vázquez-Barquero, J.L.; Cuesta Nuñez, M.J.; de la Varga, M.; Herrera Castanedo, S.; Gaite, L.; Arenal, A. The Cantabria first episode schizophrenia study: A summary of general findings. Acta Psychiatr. Scand. 1995, 91, 156–162. [Google Scholar] [CrossRef]
- Jörgensen, L.; Ahlbom, A.; Allebeck, P.; Dalman, C. The Stockholm non-affective psychoses study (snaps): The importance of including out-patient data in incidence studies. Acta Psychiatr. Scand. 2010, 121, 389–392. [Google Scholar] [CrossRef]
- Ajdacic-Gross, V.; Lauber, C.; Warnke, I.; Haker, H.; Murray, R.M.; Rössler, W. Changing incidence of psychotic disorders among the young in Zurich. Schizophr. Res. 2007, 95, 9–18. [Google Scholar] [CrossRef]
- Loganovsky, K.N.; Loganovskaja, T.K. At issue: Schizophrenia spectrum disorders in persons exposed to ionizing radiation as a result of the Chernobyl accident. Schizophr. Bull. 2000, 26, 751–773. [Google Scholar] [CrossRef] [Green Version]
- Reay, R.; Mitford, E.; McCabe, K.; Paxton, R.; Turkington, D. Incidence and diagnostic diversity in first-episode psychosis. Acta Psychiatr. Scand. 2010, 121, 315–319. [Google Scholar] [CrossRef]
- Simon, G.E.; Coleman, K.J.; Yarborough, B.J.H.; Operskalski, B.; Stewart, C.; Hunkeler, E.M.; Lynch, F.; Carrell, D.; Beck, A. First Presentation With Psychotic Symptoms in a Population-Based Sample. Psychiatr. Serv. 2017, 68, 456–461. [Google Scholar] [CrossRef] [Green Version]
- Gynther, B.; Charlson, F.; Obrecht, K.; Waller, M.; Santomauro, D.; Whiteford, H.; Hunter, E. The Epidemiology of Psychosis in Indigenous Populations in Cape York and the Torres Strait. EClinicalMedicine 2019, 10, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacono, W.G.; Beiser, M. Are males more likely than females to develop schizophrenia? Am. J. Psychiatry 1992, 149, 1070–1074. [Google Scholar] [CrossRef] [PubMed]
- Nicole, L.; Lesage, A.; Lalonde, P. Lower incidence and increased male:female ratio in schizophrenia. Br. J. Psychiatry 1992, 161, 556–557. [Google Scholar] [CrossRef] [PubMed]
- Salokangas, R.K.R. First-contact rate for schizophrenia in community psychiatric care. Consideration of the oestrogen hypothesis. Eur. Arch. Psychiatry Clin. Neurosci. 1993, 242, 337–346. [Google Scholar] [CrossRef]
- Suokas, K.; Koivisto, A.M.; Hakulinen, C.; Kaltiala, R.; Sund, R.; Lumme, S.; Kampman, O.; Pirkola, S. Association of Income With the Incidence Rates of First Psychiatric Hospital Admissions in Finland, 1996–2014. JAMA Psychiatry 2020, 77, 274–284. [Google Scholar] [CrossRef]
- De Salvia, D.; Barbato, A.; Salvo, P.; Zadro, F. Prevalence and incidence of schizophrenic disorders in Portogruaro. An Italian case register study. J. Nerv. Ment. Dis. 1993, 181, 275–282. [Google Scholar] [CrossRef]
- Preti, A.; Miotto, P. Increase in first admissions for schizophrenia and other major psychoses in Italy. Psychiatry Res. 2000, 94, 139–152. [Google Scholar] [CrossRef]
- Lasalvia, A.; Bonetto, C.; Tosato, S.; Zanatta, G.; Cristofalo, D.; Salazzari, D.; Lazzarotto, L.; Bertani, M.; Bissoli, S.; De Santi, K.; et al. First-contact incidence of psychosis in north-eastern Italy: Influence of age, gender, immigration and socioeconomic deprivation. Br. J. Psychiatry 2014, 205, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Tarricone, I.; Mimmi, S.; Paparelli, A.; Rossi, E.; Mori, E.; Panigada, S.; Carchia, G.; Bandieri, V.; Michetti, R.; Minenna, G.; et al. First-episode psychosis at the West Bologna Community Mental Health Centre: Results of an 8-year prospective study. Psychol. Med. 2012, 42, 2255–2264. [Google Scholar] [CrossRef] [Green Version]
- Park Jae-Sup, C.W.-J.; Dong-Wook, K.; Jung-Eun, S.; Seul-Bi, L.; San-Hyeop, L. Status of Treatment for First Episode Psychosis in Korea and Need for Expansion of Coverage; Report from 2017 NHIS Ilsan Hospital; National Health Insurance Service Ilsan Hospital: Goyang, Republic of Korea, 2017; p. 113. [Google Scholar]
- Orrico-Sánchez, A.; López-Lacort, M.; Munõz-Quiles, C.; Sanfélix-Gimeno, G.; Diéz-Domingo, J. Epidemiology of schizophrenia and its management over 8-years period using real-world data in Spain. BMC Psychiatry 2020, 20, 149. [Google Scholar] [CrossRef] [Green Version]
- Osby, U.; Hammar, N.; Brandt, L.; Wicks, S.; Thinsz, Z.; Ekbom, A.; Sparén, P. Time trends in first admissions for schizophrenia and paranoid psychosis in Stockholm County, Sweden. Schizophr. Res. 2001, 47, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.; Golay, P.; Boubaker, K.; Bonsack, C.; Morandi, S. Community treatment orders in Western Switzerland: A retrospective epidemiological study. Int. J. Law Psychiatry 2019, 67, 101509. [Google Scholar] [CrossRef] [PubMed]
- Brewin, J.; Cantwell, R.; Dalkin, T.; Fox, R.; Medley, I.; Glazebrook, C.; Kwiecinski, R.; Harrison, G. Incidence of schizophrenia in Nottingham: A comparison of two cohorts, 1978–80 and 1992–94. Br. J. Psychiatry 1997, 171, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Goldacre, M.; Shiwach, R.; Yeates, D. Estimating incidence and prevalence of treated psychiatric disorders from routine statistics: The example of schizophrenia in Oxfordshire. J. Epidemiol. Community Health 1994, 48, 318–322. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; DelPozo-Banos, M.; Lloyd, K.; Jones, I.; Walters, J.T.R.; Owen, M.J.; O’Donovan, M.; John, A. Area deprivation, urbanicity, severe mental illness and social drift—A population-based linkage study using routinely collected primary and secondary care data. Schizophr. Res. 2020, 220, 130–140. [Google Scholar] [CrossRef]
- Singh, S.P.; Burns, T.; Amin, S.; Jones, P.B.; Harrison, G. Acute and transient psychotic disorders: Precursors, epidemiology, course and outcome. Br. J. Psychiatry 2004, 185, 452–459. [Google Scholar] [CrossRef]
- Jensen, C.M.; Steinhausen, H.C. Time Trends in Lifetime Incidence Rates of First-Time Diagnosed Bipolar and Depressive Disorders Across 16 Years in Danish Psychiatric Hospitals: A Nationwide Study. J. Clin. Psychiatry 2016, 77, e1570–e1575. [Google Scholar] [CrossRef] [Green Version]
- Markkula, N.; Lehti, V.; Gissler, M.; Suvisaari, J. Incidence and prevalence of mental disorders among immigrants and native Finns: A register-based study. Soc. Psychiatry Psychiatr. Epidemiol. 2017, 52, 1523–1540. [Google Scholar] [CrossRef] [Green Version]
- Selten, J.P.; van Os, J.; Nolen, W.A. First admissions for mood disorders in immigrants to the Netherlands. Soc. Psychiatry Psychiatr. Epidemiol. 2003, 38, 547–550. [Google Scholar] [CrossRef]
- Carlborg, A.; Ferntoft, L.; Thuresson, M.; Bodegard, J. Population study of disease burden, management, and treatment of bipolar disorder in Sweden: A retrospective observational registry study. Bipolar Disord. 2015, 17, 76–85. [Google Scholar] [CrossRef]
- Hibbeln, J.R. Fish consumption and major depression. Lancet 1998, 351, 1213. [Google Scholar] [CrossRef]
- Noaghiul, S.; Hibbeln, J.R. Cross-national comparisons of seafood consumption and rates of bipolar disorders. Am. J. Psychiatry 2003, 160, 2222–2227. [Google Scholar] [CrossRef] [PubMed]
- Caspi, A.; Williams, B.; Kim-Cohen, J.; Craig, I.W.; Milne, B.J.; Poulton, R.; Schalkwyk, L.C.; Taylor, A.; Werts, H.; Moffitt, T.E. Moderation of breastfeeding effects on the IQ by genetic variation in fatty acid metabolism. Proc. Natl. Acad. Sci. USA 2007, 104, 18860–18865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingason, A.; Rujescu, D.; Cichon, S.; Sigurdsson, E.; Sigmundsson, T.; Pietilainen, O.P.; Buizer-Voskamp, J.E.; Strengman, E.; Francks, C.; Muglia, P.; et al. Copy number variations of chromosome 16p13.1 region associated with schizophrenia. Mol. Psychiatry 2011, 16, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horrobin, D.F. The membrane phospholipid hypothesis as a biochemical basis for the neurodevelopmental concept of schizophrenia. Schizophr. Res. 1998, 30, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Inoue, T.; Sasaki, J.; Kubo, T.; Matsuda, S.; Nakasaki, Y.; Hattori, M.; Tanaka, F.; Udagawa, O.; Kono, N.; et al. LPIAT1 regulates arachidonic acid content in phosphatidylinositol and is required for cortical lamination in mice. Mol. Biol. Cell 2012, 23, 4689–4700. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, M.; Watanabe, A.; Iwayama, Y.; Kimura, T.; Hamazaki, K.; Balan, S.; Ohba, H.; Hisano, Y.; Nozaki, Y.; Ohnishi, T.; et al. Polyunsaturated fatty acid deficiency during neurodevelopment in mice models the prodromal state of schizophrenia through epigenetic changes in nuclear receptor genes. Transl. Psychiatry 2017, 7, e1229. [Google Scholar] [CrossRef]
- Maekawa, M.; Takashima, N.; Matsumata, M.; Ikegami, S.; Kontani, M.; Hara, Y.; Kawashima, H.; Owada, Y.; Kiso, Y.; Yoshikawa, T.; et al. Arachidonic acid drives postnatal neurogenesis and elicits a beneficial effect on prepulse inhibition, a biological trait of psychiatric illnesses. PLoS ONE 2009, 4, e5085. [Google Scholar] [CrossRef] [Green Version]
- Kotani, S.; Sakaguchi, E.; Warashina, S.; Matsukawa, N.; Ishikura, Y.; Kiso, Y.; Sakakibara, M.; Yoshimoto, T.; Guo, J.; Yamashima, T. Dietary supplementation of arachidonic and docosahexaenoic acids improves cognitive dysfunction. Neurosci. Res. 2006, 56, 159–164. [Google Scholar] [CrossRef]
- Tokuda, H.; Ito, M.; Sueyasu, T.; Sasaki, H.; Morita, S.; Kaneda, Y.; Rogi, T.; Kondo, S.; Kouzaki, M.; Tsukiura, T.; et al. Effects of combining exercise with long-chain polyunsaturated fatty acid supplementation on cognitive function in the elderly: A randomised controlled trial. Sci. Rep. 2020, 10, 12906. [Google Scholar] [CrossRef]
- MacDonald-Wicks, L.; McEvoy, M.; Magennis, E.; Schofield, P.W.; Patterson, A.J.; Zacharia, K. Dietary Long-Chain Fatty Acids and Cognitive Performance in Older Australian Adults. Nutrients 2019, 11, 711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibbeln, J.R. Seafood consumption, the DHA content of mothers’ milk and prevalence rates of postpartum depression: A cross-national, ecological analysis. J. Affect. Disord. 2002, 69, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Pajak, A.; Marventano, S.; Castellano, S.; Galvano, F.; Bucolo, C.; Drago, F.; Caraci, F. Role of omega-3 fatty acids in the treatment of depressive disorders: A comprehensive meta-analysis of randomized clinical trials. PLoS ONE 2014, 9, e96905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarris, J.; Mischoulon, D.; Schweitzer, I. Omega-3 for bipolar disorder: Meta-analyses of use in mania and bipolar depression. J. Clin. Psychiatry 2012, 73, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenton, W.S.; Dickerson, F.; Boronow, J.; Hibbeln, J.R.; Knable, M. A placebo-controlled trial of omega-3 fatty acid (ethyl eicosapentaenoic acid) supplementation for residual symptoms and cognitive impairment in schizophrenia. Am. J. Psychiatry 2001, 158, 2071–2074. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotype | Sample Size | Ancestry | Women% | PUFA Concentration, % of Total Fatty Acid |
|---|---|---|---|---|
| ω-6 PUFAs a | 8631 | Europe | 51~62% | LA (19.96~24.78%), GLA (0.11~0.12%), DGLA (3.13~3.33%), AA (8.00~12.10%) |
| ω-3 PUFAs b | 8866 | Europe | 51~61% | ALA (0.14~0.44%), DHA (2.29~3.66%), EPA (0.56~0.88%), DPA (0.83~0.95%) |
| Schizophrenia c | 34,241 cases, 45,604 controls | 93.5% European 6.5% East Asian | 8~73% | Not applicable |
| Country Code | Country | Incidence of Schizophrenia | PUFA Consumption a | Economic Variable | |||||
|---|---|---|---|---|---|---|---|---|---|
| Coverage | Period | Incidence (Per 100,000) | AA (mg/Day) | DHA (mg/Day) | ω-6 LCPUFA (mg/Day) | ω-3 LCPUFA (mg/Day) | GDP Per Capita b | ||
| BRA | Brazil | Subnational | 1965–1994 | 47.2 [38] | 71.97 | 65.35 | 106.99 | 158.03 | 431.3 |
| CAN | Canada | Subnational | 1995–1998 | 33.0 [39] | 231.99 | 180.74 | 356.23 | 443.35 | 8590.8 |
| CHL | Chile | National | 2004–2017 | 13.4 [40] | 113.12 | 126.64 | 164.82 | 255.50 | 3076.6 |
| CRI | Costa Rica | National | 1979–1981 | 48.2 [41] | 59.00 | 49.29 | 84.85 | 129.73 | 355.0 |
| HRV | Croatia | National | 1980–1985 | 26.0 [42] | 100.14 | 54.09 | 165.14 | 126.67 | 4852.2 |
| DNK | Denmark | National | 1995–2008 | 9.2 [43] | 201.39 | 172.04 | 352.87 | 366.56 | 13,333.0 |
| FIN | Finland | National | 2003 | 37.8 [44] | 179.83 | 283.62 | 291.77 | 592.44 | 10,497.5 |
| DEU | Germany | National | 1987–1989 | 19.0 [45] | 187.32 | 131.59 | 325.37 | 314.24 | 2761.2 |
| ISL | Iceland | National | 1967 | 27.0 [46] | 297.87 | 561.37 | 378.27 | 1278.49 | 1418.1 |
| IND | India | Subnational | 1987–1988 | 58.5 [47] | 14.70 | 29.26 | 20.41 | 65.78 | 98.1 |
| IRL | Ireland | Subnational | 1974–1977 | 27.5 [48] | 185.71 | 120.95 | 285.16 | 355.25 | 739.3 |
| ITA | Italy | Subnational | 2008–2011 | 9.6 [49] | 220.95 | 214.39 | 338.11 | 468.64 | 18,728.4 |
| JPN | Japan | Subnational | 1979–1980 | 21.0 [50] | 89.70 | 384.11 | 120.07 | 682.04 | 563.6 |
| NLD | Netherlands | Subnational | 1981–1997 | 22.3 [51] | 164.43 | 123.68 | 267.52 | 297.9 | 3662.2 |
| NZL | New Zealand | National | 2014 | 6.3 [52] | 294.28 | 242.54 | 410.68 | 630.07 | 17,400.4 |
| NOR | Norway | Subnational | 1982–1983 | 5.5 [53] | 154.97 | 318.68 | 232.38 | 658.06 | 1721.4 |
| KOR | Republic of Korea | National | 2008–2017 | 50.0 [54] | 137.70 | 339.12 | 198.58 | 978.261 | 9057.6 |
| RUS | Russian Federation | National | 2015 | 8.0 [55] | 162.29 | 194.33 | 248.96 | 421.68 | 2665.8 |
| ESP | Spain | Subnational | 1989–1990 | 19.0 [56] | 157.49 | 263.78 | 234.63 | 525.39 | 1145.0 |
| SWE | Sweden | Subnational | 1997–2006 | 13.0 [57] | 188.92 | 229.14 | 305.55 | 503.14 | 14,272.5 |
| CHE | Switzerland | Subnational | 1977–2005 | 11.8 [58] | 188.47 | 137.66 | 308.20 | 350.49 | 8953.3 |
| UKR | Ukraine | Subnational | 1986–1997 | 11.0 [59] | 149.60 | 114.19 | 248.40 | 261.74 | 1417.9 |
| GBR | United Kingdom | Subnational | 1998–2005 | 4.1 [60] | 201.51 | 165.01 | 311.62 | 403.32 | 7937.9 |
| USA | United States | Subnational | 2007–2013 | 7.5 [61] | 274.59 | 186.38 | 406.67 | 444.37 | 23,478.6 |
| Effect Size Estimates for PUFAs a | Effect Size Estimates for Schizophrenia b | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| PUFA | SNP | Chr | Nearby Gene | Effect Allele | F Statistics | β | SE | p | ln (OR) | SE | p |
| Linoleic acid (LA, 18:2 ω-6) | rs10740118 | 10 | JMJD1C | G | 32.91 | 0.248 | 0.043 | 8.08 × 10−9 | 0.0093 | 0.0110 | 0.399 |
| rs174547 | 11 | FADS1 | C | 1218.48 | 1.474 | 0.042 | 4.98 × 10−274 | 0.0205 | 0.0114 | 0.072 | |
| rs16966952 | 16 | NTAN1, PDXDC1 | G | 62.96 | 0.351 | 0.044 | 1.23 × 10−15 | −0.0394 | 0.0117 | 0.001 | |
| γ-linolenic acid (GLA, 18:3 ω-6) | rs174547 | 11 | FADS1 | T | 253.26 | 0.016 | 0.001 | 2.29 × 10−72 | −0.0205 | 0.0114 | 0.072 |
| rs16966952 | 16 | NTAN1, PDXDC1 | G | 36.81 | 0.006 | 0.001 | 5.0 × 10−11 | −0.0394 | 0.0117 | 0.001 | |
| Dihomo-γ-linolenic acid (DGLA, 20:3 ω-6) | rs174547 | 11 | FADS1 | T | 1211.88 | −0.350 | 0.010 | 2.63 × 10−151 | −0.0205 | 0.0114 | 0.072 |
| rs16966952 | 16 | NTAN1, PDXDC1 | G | 119.70 | 0.220 | 0.020 | 7.55 × 10−65 | −0.0394 | 0.0117 | 0.001 | |
| Arachidonic acid (AA, 20:4 ω-6) | rs174547 | 11 | FADS1 | T | 4526.15 | 1.691 | 0.025 | 3.00 × 10−971 | −0.0205 | 0.0114 | 0.072 |
| rs16966952 | 16 | NTAN1, PDXDC1 | G | 40.77 | 0.199 | 0.031 | 2.43 × 10−10 | −0.0394 | 0.0117 | 0.001 | |
| α-linolenic acid (ALA, 18:3 ω-3) | rs174547 | 11 | FADS1 | C | 256.00 | 0.016 | 0.001 | 3.47 × 10−64 | 0.0205 | 0.0114 | 0.072 |
| Eicosapentaenoic acid (EPA, 20:5ω3) | rs3798713 | 6 | ELOVL2 | C | 49.00 | 0.035 | 0.005 | 1.93 × 10−12 | 0.0081 | 0.0110 | 0.459 |
| rs174538 | 11 | FADS1 | G | 275.56 | 0.083 | 0.005 | 5.37 × 10−58 | −0.0211 | 0.0117 | 0.072 | |
| Docosapentaenoic acid (DPA, 22:5 ω-3) | rs780094 | 2 | GCKR | T | 32.11 | 0.017 | 0.003 | 9.04 × 10−9 | 0.0007 | 0.0111 | 0.948 |
| rs3734398 | 6 | ELOVL2 | C | 177.78 | 0.040 | 0.003 | 9.61 × 10−44 | 0.0088 | 0.0110 | 0.424 | |
| rs174547 | 11 | FADS1 | T | 625.00 | 0.075 | 0.003 | 3.79 × 10−154 | −0.0205 | 0.0114 | 0.072 | |
| Docosahexaenoic acid (DHA, 22:6 ω-3) | rs2236212 | 6 | ELOVL2 | G | 65.15 | 0.113 | 0.014 | 1.26 × 10−15 | −0.0085 | 0.0110 | 0.439 |
| Number of SNP Instruments (Related to PUFA Group) a | Number of SNPs in Analysis | F Statistic | Result in IVW Method a,b | ||
|---|---|---|---|---|---|
| OR (95% CI) | p | ||||
| Arachidonic acid (AA, 20:4 ω-6) | 3 | 8 | 85.33 | 0.981 (0.976–0.986) | 0.009 |
| Eicosapentaenoic acid (EPA, 20:5 ω-3) | 6 | 8 | 30.67 | 1.359 (0.702–2.633) | 0.090 |
| Linoleic acid (LA, 18:2 ω-6) | 3 | 8 | 6.46 | 1.005 (0.957–1.055) | 0.805 |
| α-linolenic acid (ALA, 18:3 ω-3) | 6 | 8 | 6.02 | 3.747 (0.019–728.509) | 0.512 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Hu, X.; Wang, D.; Jiang, J.; Li, M.; Qing, Y.; Yang, X.; Zhang, J.; Zhang, Y.; Wan, C. Association between Arachidonic Acid and the Risk of Schizophrenia: A Cross-National Study and Mendelian Randomization Analysis. Nutrients 2023, 15, 1195. https://doi.org/10.3390/nu15051195
Gao Y, Hu X, Wang D, Jiang J, Li M, Qing Y, Yang X, Zhang J, Zhang Y, Wan C. Association between Arachidonic Acid and the Risk of Schizophrenia: A Cross-National Study and Mendelian Randomization Analysis. Nutrients. 2023; 15(5):1195. https://doi.org/10.3390/nu15051195
Chicago/Turabian StyleGao, Yan, Xiaowen Hu, Dandan Wang, Jie Jiang, Minghui Li, Ying Qing, Xuhan Yang, Juan Zhang, Yue Zhang, and Chunling Wan. 2023. "Association between Arachidonic Acid and the Risk of Schizophrenia: A Cross-National Study and Mendelian Randomization Analysis" Nutrients 15, no. 5: 1195. https://doi.org/10.3390/nu15051195
APA StyleGao, Y., Hu, X., Wang, D., Jiang, J., Li, M., Qing, Y., Yang, X., Zhang, J., Zhang, Y., & Wan, C. (2023). Association between Arachidonic Acid and the Risk of Schizophrenia: A Cross-National Study and Mendelian Randomization Analysis. Nutrients, 15(5), 1195. https://doi.org/10.3390/nu15051195