
Effects of Green and Gold Kiwifruit Varieties on Antioxidant Neuroprotective Potential in Pigs as a Model for Human Adults

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
- Soda bread only.
- Soda bread + gold KF (Actinidia chinensis cv. ‘Hort16A’).
- Soda bread + green KF (Actinidia deliciosa cv. ‘Hayward’).
2.2. Euthanasia and Tissue Collection
2.3. Biochemcial Measures
2.3.1. Plasma Oxidative Stress Measures
2.3.2. Plasma and Brain Tissue Antioxidant Measures
Oxidative Potential Assay (OPA)
Ferric-Reducing Antioxidant Power (FRAP)
Oxygen Radical Absorbance Capacity (ORAC)
Vitamin C (Ascorbate) Concentration
Catalase Activity
Superoxide Dismutase Activity
Western Blot Analysis of Antioxidant Enzymes
AChE Activity
2.4. Data Analysis
3. Results
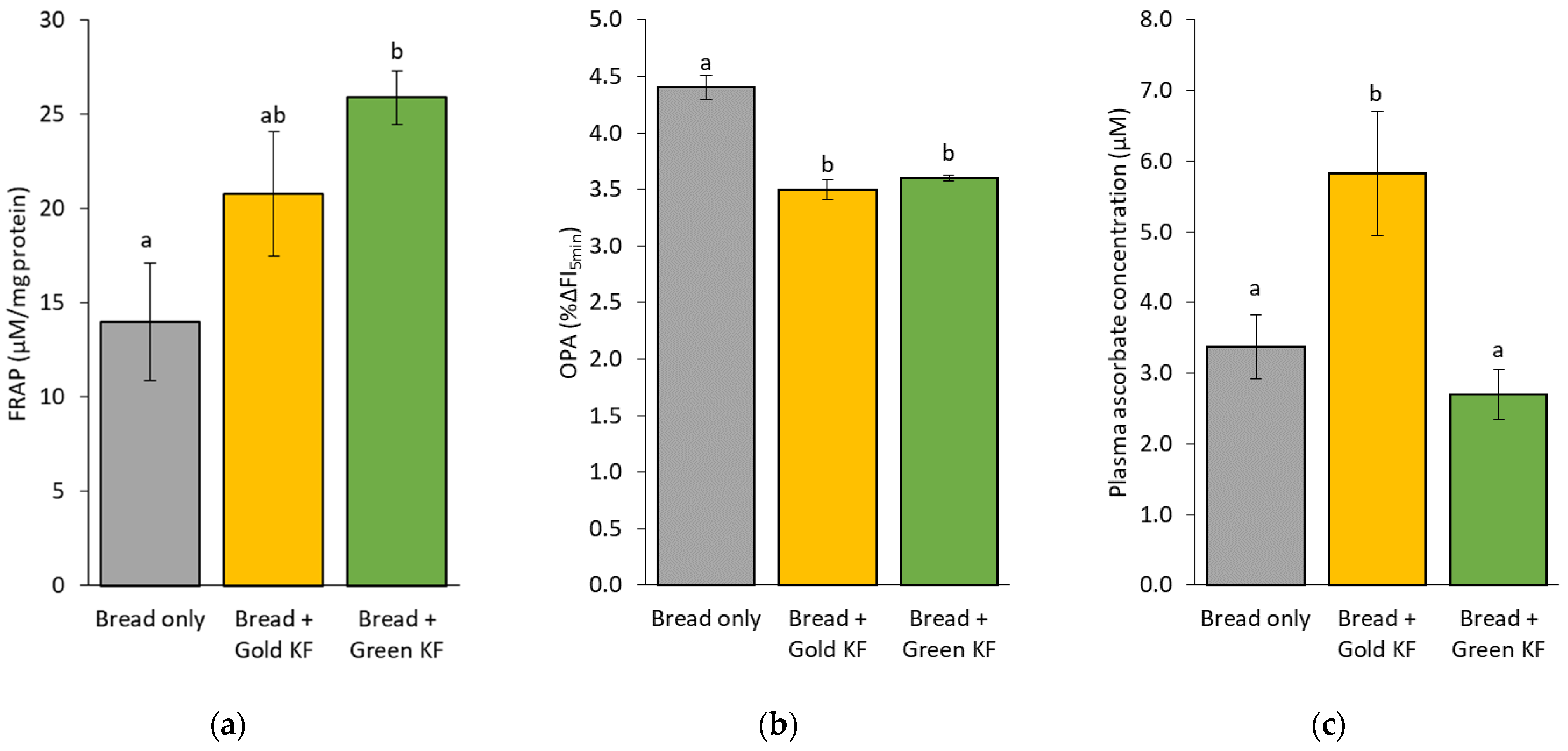
3.1. Plasma Oxidative and Antioxidant Markers
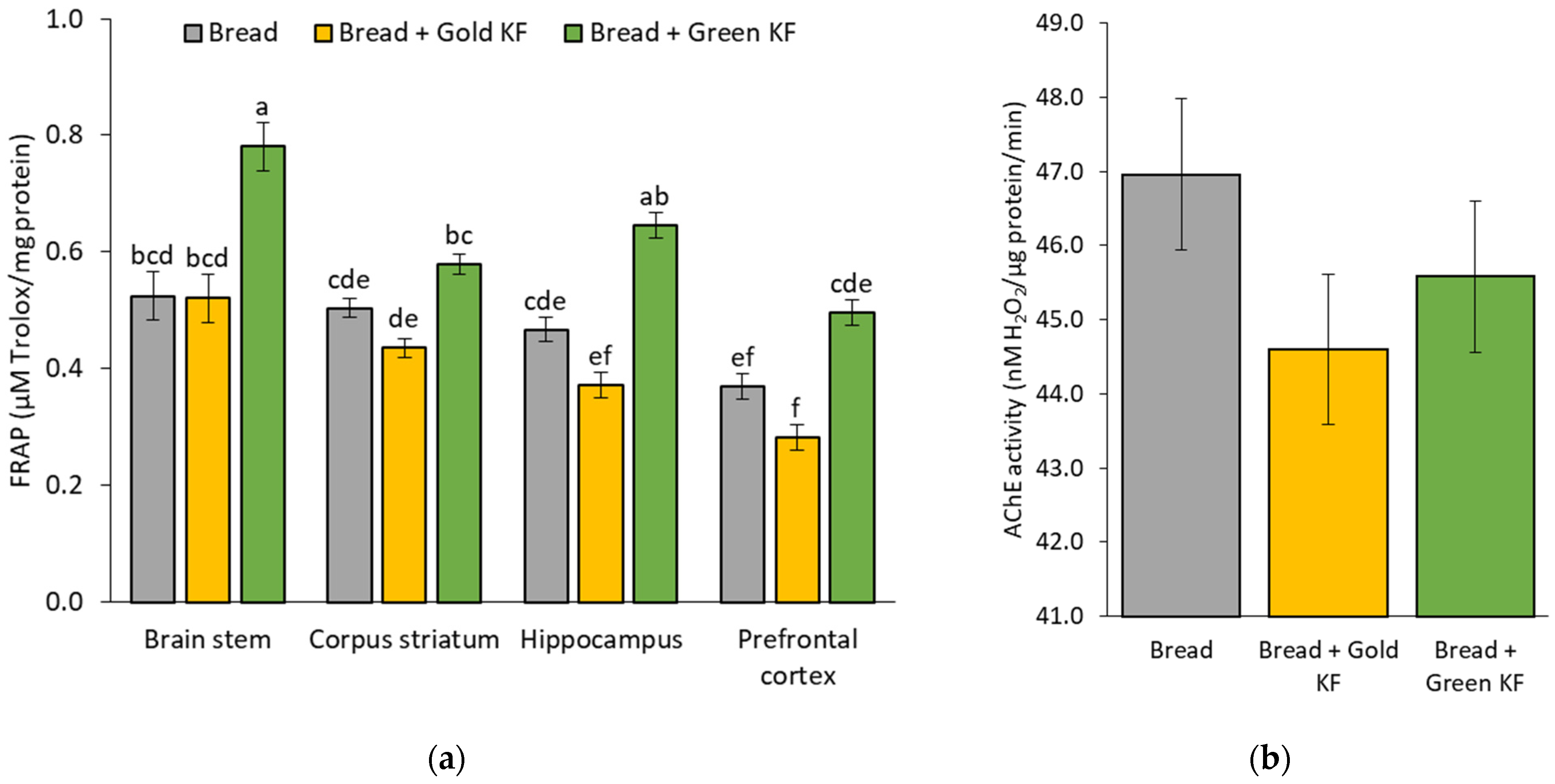
3.2. Antioxidant Enzyme Activity and Capacity in Different Brain Regions
3.3. Antioxidant Enzyme Activity and Capacity after Kiwifruit Intake
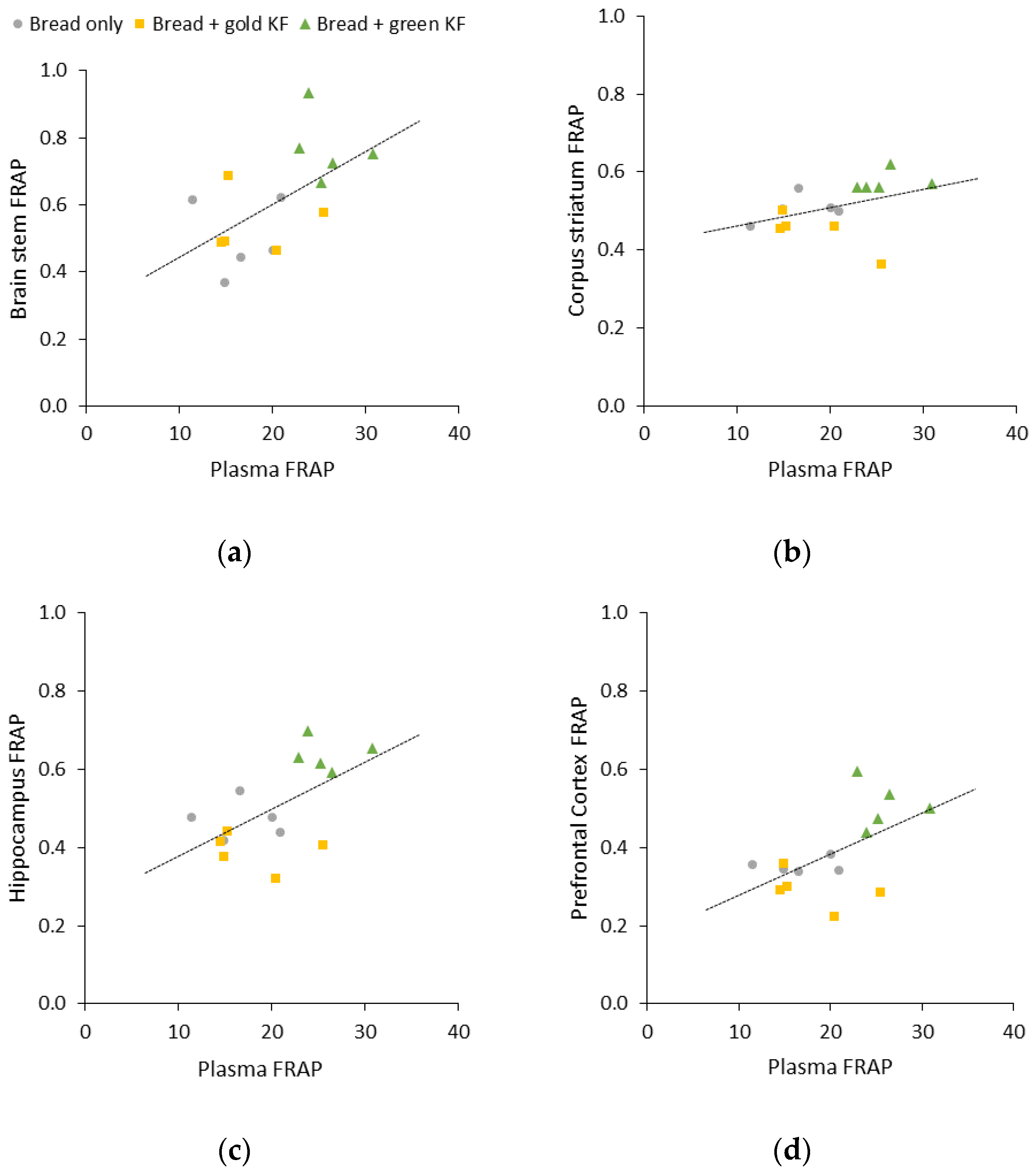
3.4. Correlations between Plasma and Brain Regional FRAP
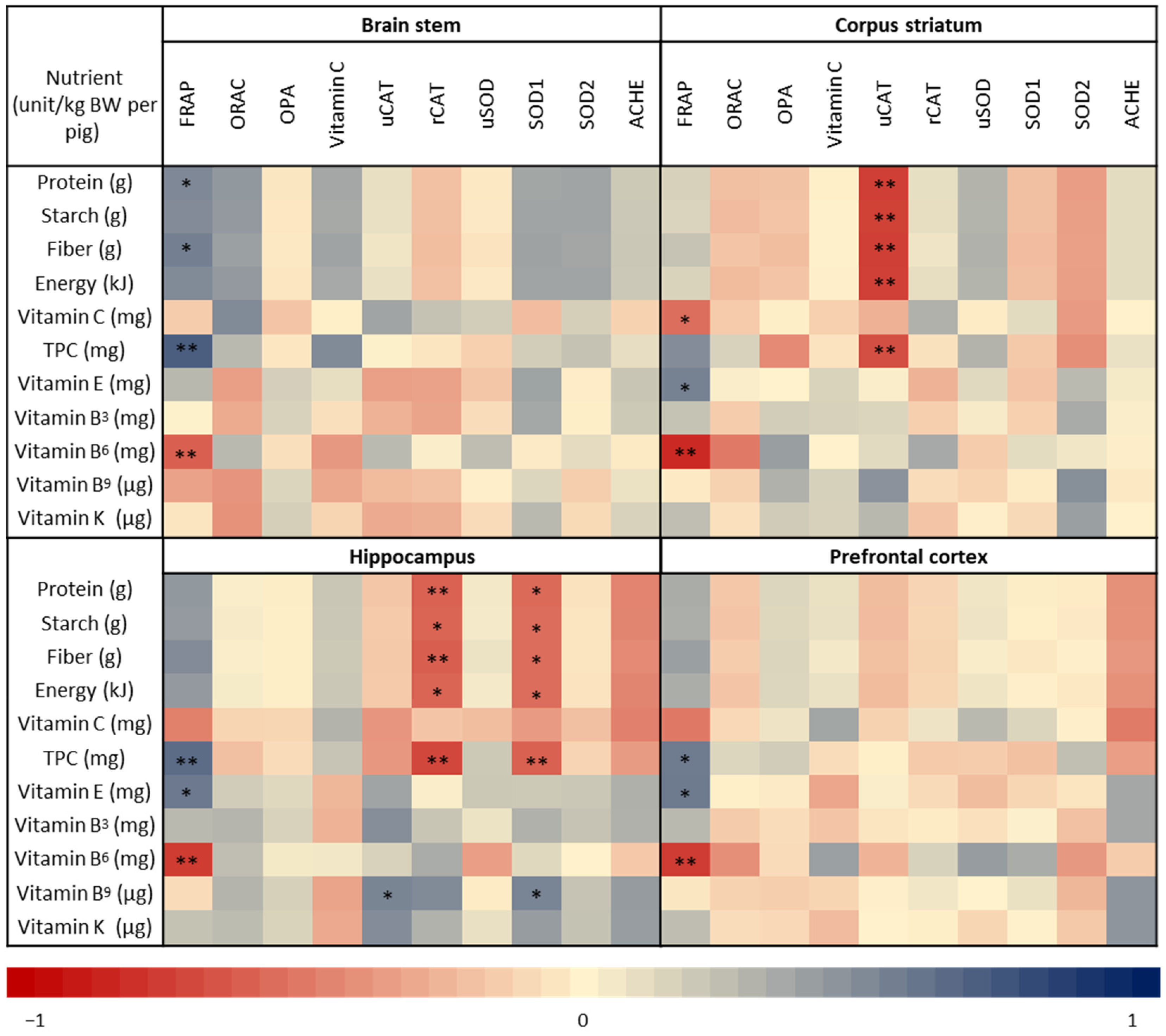
3.5. Correlation between Nutrient Intake and Antioxidant Capcity in the Brain Regions
4. Discussion
4.1. Effects of Kiwifruit on Peripheral Antioxidant Capacity
4.2. Effects of Kiwifruit on Brain Antioxidant Capacity
4.3. Correlations between Peripheral and Brain Antioxidant Capacity
4.4. Assocations between Kiwifruit Composition and Total Antioxidant Capacity
4.5. Strengths, Limitations, and Future Studies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Islam, M.T. Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol. Res. 2017, 39, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.V.; Balachandran, B. Role of oxidative stress and antioxidants in neurodegenerative diseases. Nutr. Neurosci. 2002, 5, 291–309. [Google Scholar] [CrossRef] [PubMed]
- Godos, J.; Grosso, G. Dietary Antioxidants and Brain Health: Focus on Cognitive and Affective Disorders. Antioxidants 2021, 10, 1659. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pinilla, F. Brain foods: The effects of nutrients on brain function. Nat. Rev. Neurosci. 2008, 9, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Cha, M.; Lee, B.H. Neuroprotective effect of antioxidants in the brain. Int. J. Mol. Sci. 2020, 21, 7152. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Islam, M.R.; Ahmed, I.; Moktadir, A.A.; Nahar, Z.; Islam, M.S.; Shahid, S.F.B.; Islam, S.N.; Islam, M.S.; Hasnat, A. Elevated serum levels of malondialdehyde and cortisol are associated with major depressive disorder: A case-control study. SAGE Open Med. 2018, 6, 2050312118773953. [Google Scholar] [CrossRef] [PubMed]
- Colovic, M.B.; Krstic, D.Z.; Lazarevic-Pasti, T.D.; Bondzic, A.M.; Vasic, V.M. Acetylcholinesterase inhibitors: Pharmacology and toxicology. Curr. Neuropharmacol. 2013, 11, 315–335. [Google Scholar] [CrossRef]
- Carr, A.C.; Bozonet, S.M.; Pullar, J.M.; Vissers, M.C. Mood improvement in young adult males following supplementation with gold kiwifruit, a high-vitamin C food. J. Nutr. Sci. 2013, 2, e24. [Google Scholar] [CrossRef] [PubMed]
- Conner, T.S.; Fletcher, B.D.; Pullar, J.M.; Spencer, E.; Mainvil, L.A.; Vissers, M.C.M. KiwiC for Vitality: Results of a Randomized Placebo-Controlled Trial Testing the Effects of Kiwifruit or Vitamin C Tablets on Vitality in Adults with Low Vitamin C Levels. Nutrients 2020, 12, 2898. [Google Scholar] [CrossRef]
- Lin, H.H.; Tsai, P.S.; Fang, S.C.; Liu, J.F. Effect of kiwifruit consumption on sleep quality in adults with sleep problems. Asia Pac. J. Clin. Nutr. 2011, 20, 169–174. [Google Scholar]
- Hunter, D.C.; Skinner, M.A.; Wolber, F.M.; Booth, C.L.; Loh, J.M.; Wohlers, M.; Stevenson, L.M.; Kruger, M.C. Consumption of gold kiwifruit reduces severity and duration of selected upper respiratory tract infection symptoms and increases plasma vitamin C concentration in healthy older adults. Br. J. Nutr. 2012, 108, 1235–1245. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Gu, L.; Wu, X.; Jacob, R.A.; Sotoudeh, G.; Kader, A.A.; Cook, R.A. Plasma antioxidant capacity changes following a meal as a measure of the ability of a food to alter in vivo antioxidant status. J. Am. Coll. Nutr. 2007, 26, 170–181. [Google Scholar] [CrossRef]
- Duttaroy, A.K.; Jorgensen, A. Effects of kiwi fruit consumption on platelet aggregation and plasma lipids in healthy human volunteers. Platelets 2004, 15, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.S.; Kang, J.Y.; Kang, J.E.; Park, S.K.; Kim, J.M.; Kim, C.W.; Oh, S.I.; Lee, U.; Kim, D.O.; Heo, H.J. Pentacyclic triterpenoid-rich fraction of the Hardy kiwi (Actinidia arguta) improves brain dysfunction in high fat diet-induced obese mice. Sci. Rep. 2020, 10, 5788. [Google Scholar] [CrossRef]
- Xue, W.Z.; Yang, Q.Q.; Chen, Y.; Zou, R.X.; Xing, D.; Xu, Y.; Liu, Y.S.; Wang, H.L. Kiwifruit Alleviates Learning and Memory Deficits Induced by Pb through Antioxidation and Inhibition of Microglia Activation In Vitro and In Vivo. Oxid. Med. Cell Longev. 2017, 2017, 5645324. [Google Scholar] [CrossRef]
- Cho, C.H.; Jung, Y.S.; Kim, J.M.; Nam, T.G.; Lee, S.-H.; Cho, H.S.; Song, M.C.; Heo, H.J.; Kim, D.-O. Neuroprotective effects of Actinidia eriantha cv. Bidan kiwifruit on amyloid beta-induced neuronal damages in PC-12 cells and ICR mice. J. Funct. Foods 2021, 79, 104398. [Google Scholar] [CrossRef]
- Jeong, H.R.; Kim, K.J.; Lee, S.G.; Cho, H.S.; Cho, Y.S.; Kim, D.O. Phenolic Profiles of Hardy Kiwifruits and Their Neuroprotective Effects on PC-12 and SH-SY5Y Cells against Oxidative Stress. J. Microbiol. Biotechnol. 2020, 30, 912–919. [Google Scholar] [CrossRef]
- Roura, E.; Koopmans, S.J.; Lalles, J.P.; Le Huerou-Luron, I.; de Jager, N.; Schuurman, T.; Val-Laillet, D. Critical review evaluating the pig as a model for human nutritional physiology. Nutr. Res. Rev. 2016, 29, 60–90. [Google Scholar] [CrossRef]
- Boland, M. Chapter Four—Kiwifruit Proteins and Enzymes: Actinidin and Other Significant Proteins. In Advances in Food and Nutrition Research; Boland, M., Moughan, P.J., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 68, pp. 59–80. [Google Scholar]
- Ciardiello, M.A.; Meleleo, D.; Saviano, G.; Crescenzo, R.; Carratore, V.; Camardella, L.; Gallucci, E.; Micelli, S.; Tancredi, T.; Picone, D.; et al. Kissper, a kiwi fruit peptide with channel-like activity: Structural and functional features. J. Pept. Sci. 2008, 14, 742–754. [Google Scholar] [CrossRef] [PubMed]
- Ciacci, C.; Russo, I.; Bucci, C.; Iovino, P.; Pellegrini, L.; Giangrieco, I.; Tamburrini, M.; Ciardiello, M.A. The kiwi fruit peptide kissper displays anti-inflammatory and anti-oxidant effects in in-vitro and ex-vivo human intestinal models. Clin. Exp. Immunol. 2014, 175, 476–484. [Google Scholar] [CrossRef]
- Jayawardana, I.A. Effect of Kiwifruit Actinidin on the Digestion of Gluten Proteins: A Thesis Presented in Partial Fulfilment of the Requirements for the Degree of Doctor of Philosophy in Nutritional Sciences at Massey University. Doctoral Dissertation, Massey University, Palmerston North, New Zealand, 2022. [Google Scholar]
- Montoya, C.A.; Rutherfurd, S.M.; Olson, T.D.; Purba, A.S.; Drummond, L.N.; Boland, M.J.; Moughan, P.J. Actinidin from kiwifruit (Actinidia deliciosa cv. Hayward) increases the digestion and rate of gastric emptying of meat proteins in the growing pig. Br. J. Nutr. 2014, 111, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Council, N.R. Nutrient Requirements of Swine; The National Academies Press: Washington, DC, USA, 2012; p. 420. [Google Scholar]
- Zhang, T.; Wang, W.; Huang, J.; Liu, X.; Zhang, H.; Zhang, N. Metabolomic investigation of regional brain tissue dysfunctions induced by global cerebral ischemia. BMC Neurosci. 2016, 17, 25. [Google Scholar] [CrossRef] [PubMed]
- Karatepe, M. Simultaneous determination of ascorbic acid and free malondialdehyde in human serum by HPLC-UV. LC-GC N. Am. 2004, 22, S104. [Google Scholar]
- Levine, R.L.; Williams, J.A.; Stadtman, E.P.; Shacter, E. [37] Carbonyl assays for determination of oxidatively modified proteins. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1994; Volume 233, pp. 346–357. [Google Scholar]
- Wang, H.; Joseph, J.A. Quantifying cellular oxidative stress by dichlorofluorescein assay using microplate reader. Free Radic. Biol. Med. 1999, 27, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Hoang, H.; Gu, L.; Wu, X.; Bacchiocca, M.; Howard, L.; Hampsch-Woodill, M.; Huang, D.; Ou, B.; Jacob, R. Assays for hydrophilic and lipophilic antioxidant capacity (oxygen radical absorbance capacity (ORAC(FL))) of plasma and other biological and food samples. J. Agric. Food Chem. 2003, 51, 3273–3279. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.C.; Pullar, J.M.; Moran, S.; Vissers, M.C. Bioavailability of vitamin C from kiwifruit in non-smoking males: Determination of ‘healthy’ and ‘optimal’ intakes. J. Nutr. Sci. 2012, 1, e14. [Google Scholar] [CrossRef] [PubMed]
- Peskin, A.V.; Winterbourn, C.C. A microtiter plate assay for superoxide dismutase using a water-soluble tetrazolium salt (WST-1). Clin. Chim. Acta 2000, 293, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Khamis, H. Measures of Association: How to Choose? J. Diagn. Med. Sonogr. 2008, 24, 155–162. [Google Scholar] [CrossRef]
- Lv, J.M.; Gouda, M.; Ye, X.Q.; Shao, Z.P.; Chen, J.C. Evaluation of Proanthocyanidins from Kiwi Leaves (Actinidia chinensis) against Caco-2 Cells Oxidative Stress through Nrf2-ARE Signaling Pathway. Antioxidants 2022, 11, 1367. [Google Scholar] [CrossRef]
- Peng, Y.; Cordiner, S.B.; Sawyer, G.M.; McGhie, T.K.; Espley, R.V.; Allan, A.C.; Hurst, R.D. Kiwifruit with high anthocyanin content modulates NF-κB activation and reduces CCL11 secretion in human alveolar epithelial cells. J. Funct. Foods 2020, 65, 103734. [Google Scholar] [CrossRef]
- Walton, M.C.; Lentle, R.G.; Reynolds, G.W.; Kruger, M.C.; McGhie, T.K. Anthocyanin absorption and antioxidant status in pigs. J. Agric. Food Chem. 2006, 54, 7940–7946. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Mehta, S.; Starck, C.; Wong, M.; O’Brien, W.J.; Haswell, C.; McNabb, W.; Rutherfurd-Markwick, K.; Nasef, N.A. Effect of SunGold(TM) Kiwifruit and Vitamin C Consumption on Ameliorating Exercise-Induced Stress Response in Women. Mol. Nutr. Food Res. 2021, 65, e2001219. [Google Scholar] [CrossRef]
- Cao, G.; Russell, R.M.; Lischner, N.; Prior, R.L. Serum antioxidant capacity is increased by consumption of strawberries, spinach, red wine or vitamin C in elderly women. J. Nutr. 1998, 128, 2383–2390. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Michaelis, E.K. Selective neuronal vulnerability to oxidative stress in the brain. Front. Aging Neurosci. 2010, 2, 12. [Google Scholar] [CrossRef] [PubMed]
- Salim, S. Oxidative Stress and the Central Nervous System. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef]
- Strauss, V.; Rey Moreno, M.C.; Vogt, J.; Dammann, M.; Schneider, S.; Groters, S.; van Ravenzwaay, B. Acetylcholinesterase measurement in various brain regions and muscles of juvenile, adolescent, and adult rats. Toxicol. Mech. Methods 2017, 27, 666–676. [Google Scholar] [CrossRef]
- Bozonet, S.M.; Carr, A.C.; Pullar, J.M.; Vissers, M.C. Enhanced human neutrophil vitamin C status, chemotaxis and oxidant generation following dietary supplementation with vitamin C-rich SunGold kiwifruit. Nutrients 2015, 7, 2574–2588. [Google Scholar] [CrossRef]
- Carr, A.C.; Bozonet, S.M.; Pullar, J.M.; Simcock, J.W.; Vissers, M.C. A randomized steady-state bioavailability study of synthetic versus natural (kiwifruit-derived) vitamin C. Nutrients 2013, 5, 3684–3695. [Google Scholar] [CrossRef]
- Wilson, R.; Willis, J.; Gearry, R.B.; Hughes, A.; Lawley, B.; Skidmore, P.; Frampton, C.; Fleming, E.; Anderson, A.; Jones, L.; et al. SunGold Kiwifruit Supplementation of Individuals with Prediabetes Alters Gut Microbiota and Improves Vitamin C Status, Anthropometric and Clinical Markers. Nutrients 2018, 10, 895. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.H.; Liu, J.F. Effects of kiwifruit consumption on serum lipid profiles and antioxidative status in hyperlipidemic subjects. Int. J. Food Sci. Nutr. 2009, 60, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Graziani, F.; Discepoli, N.; Gennai, S.; Karapetsa, D.; Nisi, M.; Bianchi, L.; Rosema, N.A.M.; Van der Velden, U. The effect of twice daily kiwifruit consumption on periodontal and systemic conditions before and after treatment: A randomized clinical trial. J. Periodontol. 2018, 89, 285–293. [Google Scholar] [CrossRef]
- Mahan, D.C.; Ching, S.; Dabrowski, K. Developmental aspects and factors influencing the synthesis and status of ascorbic Acid in the pig. Annu. Rev. Nutr. 2004, 24, 79–103. [Google Scholar] [CrossRef] [PubMed]
- Travica, N.; Ried, K.; Sali, A.; Scholey, A.; Hudson, I.; Pipingas, A. Vitamin C Status and Cognitive Function: A Systematic Review. Nutrients 2017, 9, 960. [Google Scholar] [CrossRef]
- Mefford, I.N.; Oke, A.F.; Adams, R.N. Regional distribution of ascorbate in human brain. Brain Res. 1981, 212, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Harrison, F.E.; May, J.M. Vitamin C function in the brain: Vital role of the ascorbate transporter SVCT2. Free Radic. Biol. Med. 2009, 46, 719–730. [Google Scholar] [CrossRef]
- Mun, G.H.; Kim, M.J.; Lee, J.H.; Kim, H.J.; Chung, Y.H.; Chung, Y.B.; Kang, J.S.; Hwang, Y.I.; Oh, S.H.; Kim, J.G.; et al. Immunohistochemical study of the distribution of sodium-dependent vitamin C transporters in adult rat brain. J. Neurosci. Res. 2006, 83, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.A.; Reynolds, J.N. Spontaneous firing and evoked pauses in the tonically active cholinergic interneurons of the striatum. Neuroscience 2011, 198, 27–43. [Google Scholar] [CrossRef]
- Ahmadi, N.; Hosseini, M.J.; Rostamizadeh, K.; Anoush, M. Investigation of therapeutic effect of curcumin alpha and beta glucoside anomers against Alzheimer’s disease by the nose to brain drug delivery. Brain Res. 2021, 1766, 147517. [Google Scholar] [CrossRef]
- Kim, H.K.; Kim, M.; Kim, S.; Kim, M.; Chung, J.H. Effects of green tea polyphenol on cognitive and acetylcholinesterase activities. Biosci. Biotechnol. Biochem. 2004, 68, 1977–1979. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.J.; Oh, C.-S.; Park, Y.-D.; Eom, S.H.; Kim, D.-O.; Kim, U.-J.; Cho, Y.-S. Physiological components of kiwifruits with in vitro antioxidant and acetylcholinesterase inhibitory activities. Food Sci. Biotechnol. 2014, 23, 943–949. [Google Scholar] [CrossRef]
- Haytowitz, D.B.; Wu, X.; Bhagwat, S. USDA Database for the Flavonoid Content of Selected Foods, Release 3.3. U.S. Department of Agriculture, Agricultural Research Service. Available online: http://www.ars.usda.gov/nutrientdata/flav (accessed on 2 February 2024).
- Jabir, N.R.; Khan, F.R.; Tabrez, S. Cholinesterase targeting by polyphenols: A therapeutic approach for the treatment of Alzheimer’s disease. CNS Neurosci. Ther. 2018, 24, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.; Marya; Amin, S.; Kamal, M.A.; Patel, S. Flavonoids as acetylcholinesterase inhibitors: Current therapeutic standing and future prospects. Biomed. Pharmacother. 2018, 101, 860–870. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Devaki, M. The ferric reducing/antioxidant power (FRAP) assay for non-enzymatic antioxidant capacity: Concepts, procedures, limitations and applications. In Measurement of Antioxidant Activity & Capacity; Wiley: Hoboken, NJ, USA, 2017; pp. 77–106. [Google Scholar]
- Moughan, P.; Cranwell, P.; Darragh, A.; Rowan, A. The domestic pig as a model animal for studying digestion in humans. Publ.-Eur. Assoc. Anim. Prod. 1994, 80, 389. [Google Scholar]
- Lind, N.M.; Moustgaard, A.; Jelsing, J.; Vajta, G.; Cumming, P.; Hansen, A.K. The use of pigs in neuroscience: Modeling brain disorders. Neurosci. Biobehav. Rev. 2007, 31, 728–751. [Google Scholar] [CrossRef]
- Simchick, G.; Shen, A.; Campbell, B.; Park, H.J.; West, F.D.; Zhao, Q. Pig Brains Have Homologous Resting-State Networks with Human Brains. Brain Connect. 2019, 9, 566–579. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.N.; Ipsen, D.H.; Schou-Pedersen, A.M.; Lykkesfeldt, J.; Tveden-Nyborg, P. Long term Westernized diet leads to region-specific changes in brain signaling mechanisms. Neurosci. Lett. 2018, 676, 85–91. [Google Scholar] [CrossRef]
- Yeste, N.; Perez-Valle, J.; Heras-Molina, A.; Pesantez-Pacheco, J.L.; Porrini, E.; Gonzalez-Bulnes, A.; Bassols, A. A High-Fat Diet Modifies Brain Neurotransmitter Profile and Hippocampal Proteome and Morphology in an IUGR Pig Model. Nutrients 2022, 14, 3440. [Google Scholar] [CrossRef]
- Menneson, S.; Menicot, S.; Ferret-Bernard, S.; Guerin, S.; Rome, V.; Le Normand, L.; Randuineau, G.; Gambarota, G.; Noirot, V.; Etienne, P.; et al. Validation of a Psychosocial Chronic Stress Model in the Pig Using a Multidisciplinary Approach at the Gut-Brain and Behavior Levels. Front. Behav. Neurosci. 2019, 13, 161. [Google Scholar] [CrossRef]
- Kornum, B.R.; Knudsen, G.M. Cognitive testing of pigs (Sus scrofa) in translational biobehavioral research. Neurosci. Biobehav. Rev. 2011, 35, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Liu, H.; Zhao, T.; Meng, C.; Liu, Z.; Liu, X. Bioactive compounds and in vitro antioxidant activities of peel, flesh and seed powder of kiwi fruit. Int. J. Food Sci. Technol. 2018, 53, 2239–2245. [Google Scholar] [CrossRef]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.W.; Haskell-Ramsay, C.F.; Kennedy, D.O.; Cooney, J.M.; Trower, T.; Scheepens, A. Acute supplementation with blackcurrant extracts modulates cognitive functioning and inhibits monoamine oxidase-B in healthy young adults. J. Funct. Foods 2015, 17, 524–539. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition (Unit/kg BW per Pig) | Bread Only | Bread + Gold KF | Bread + Green KF |
|---|---|---|---|
| Dry matter (g) | 40.81 | 40.61 | 41.68 |
| Protein (g) | 2.80 | 2.91 | 3.00 |
| Starch (g) | 14.34 | 14.82 | 15.23 |
| Fiber (g) | 2.09 | 2.15 | 2.24 |
| Gross energy (kJ) | 493.91 | 510.51 | 525.12 |
| Vitamin C (mg) | 0.27 | 13.27 | 4.69 |
| Vitamin E (mg) | 3.38 | 2.87 | 3.30 |
| Vitamin K (μg) | 0.18 | 0.15 | 0.17 |
| Vitamin B3 (mg) | 1.64 | 1.48 | 1.56 |
| Vitamin B6 (mg) | 0.30 | 0.31 | 0.28 |
| Vitamin B9 (μg) | 23.06 | 19.16 | 19.44 |
| Total polyphenol content (mg GAE) | 0.22 | 0.63 | 1.17 |
| GAE: gallic acid equivalents |
| Measure | Treatment | Brain Stem | Corpus Striatum | Hippocampus | Prefrontal Cortex | Factors | F | p |
|---|---|---|---|---|---|---|---|---|
| FRAP (µM/mg protein) | Bread only | 0.52 ± 0.04 bcd | 0.50 ± 0.02 cde | 0.47 ± 0.02 cde | 0.37 ± 0.02 ef | Treatment | 52.52 | <0.001 |
| Bread + gold KF | 0.52 ± 0.04 bcd | 0.44 ± 0.02 de | 0.37 ± 0.02 ef | 0.28 ± 0.02 f | Brain region | 49.48 | <0.001 | |
| Bread + green KF | 0.78 ± 0.04 a | 0.58 ± 0.02 bc | 0.65 ± 0.02 ab | 0.50 ± 0.02 cde | Treatment × regions | 4.33 | 0.010 | |
| ORAC (µM/mg protein) | Bread only | 34.01 ± 2.94 | 23.40 ± 2.18 | 29.86 ± 2.94 | 26.01 ± 2.57 | Treatment | 0.27 | 0.764 |
| Bread + gold KF | 43.64 ± 2.94 | 22.21 ± 2.18 | 27.37 ± 2.94 | 25.20 ± 2.57 | Brain region | 27.03 | <0.001 | |
| Bread + green KF | 39.73 ± 2.94 | 25.50 ± 2.18 | 26.34 ± 2.94 | 27.98 ± 2.57 | Treatment × regions | 1.72 | 0.184 | |
| OPA (%ΔFI5min/mg protein) | Bread only | −0.47 ± 2.26 | 8.76 ± 3.72 | 10.19 ± 3.13 | 24.49 ± 8.97 | Treatment | 0.01 | 0.995 |
| Bread + gold KF | −2.65 ± 2.26 | 6.76 ± 3.72 | 8.13 ± 3.13 | 29.72 ± 8.97 | Brain region | 41.07 | <0.001 | |
| Bread + green KF | −1.14 ± 2.26 | −0.72 ± 3.72 | 8.59 ± 3.13 | 34.26 ± 8.97 | Treatment × regions | 0.73 | 0.631 | |
| Ascorbate (nmol/mg tissue) | Bread only | 0.64 ± 0.02 | 0.80 ± 0.04 | 1.25 ± 0.03 | 1.00 ± 0.07 | Treatment | 1.00 | 0.392 |
| Bread + gold KF | 0.65 ± 0.02 | 0.77 ± 0.04 | 1.31 ± 0.03 | 1.12 ± 0.07 | Brain region | 379.62 | <0.001 | |
| Bread + green KF | 0.69 ± 0.02 | 0.79 ± 0.04 | 1.30 ± 0.03 | 0.97 ± 0.07 | Treatment × regions | 1.25 | 0.338 | |
| Catalase activity (U/mg protein) | Bread only | 15.53 ± 0.73 | 13.08 ± 0.37 | 13.92 ± 0.75 | 13.07 ± 0.78 | Treatment | 1.11 | 0.354 |
| Bread + gold KF | 17.04 ± 0.73 | 12.10 ± 0.37 | 11.98 ± 0.75 | 12.62 ± 0.78 | Brain region | 30.54 | <0.001 | |
| Bread + green KF | 15.72 ± 0.73 | 11.30 ± 0.37 | 12.10 ± 0.75 | 13.04 ± 0.78 | Treatment × regions | 2.10 | 0.114 | |
| Catalase (relative expression) | Bread only | 1.02 ± 0.07 | 0.98 ± 0.11 | 1.11 ± 0.06 | 1.01 ± 0.09 | Treatment | 2.53 | 0.113 |
| Bread + gold KF | 1.11 ± 0.07 | 1.17 ± 0.11 | 0.97 ± 0.06 | 1.02 ± 0.09 | Brain region | 0.78 | 0.525 | |
| Bread + green KF | 1.01 ± 0.07 | 0.95 ± 0.11 | 0.79 ± 0.06 | 0.93 ± 0.09 | Treatment × regions | 1.26 | 0.331 | |
| SOD activity (U/mg protein) | Bread only | 101.5 ± 18.3 | 327.1 ± 37.9 | 664.2 ± 41.6 | 458.8 ± 89.1 | Treatment | 0.30 | 0.746 |
| Bread + gold KF | 115.4 ± 18.3 | 334.6 ± 37.9 | 623.6 ± 41.6 | 578.9 ± 89.1 | Brain region | 211.87 | <0.001 | |
| Bread + green KF | 88.3 ± 18.3 | 392.7 ± 37.9 | 707.3 ± 41.6 | 394.5 ± 89.1 | Treatment × regions | 0.92 | 0.510 | |
| SOD1 (relative expression) | Bread only | 1.10 ± 0.30 | 0.96 ± 0.10 | 1.21 ± 0.07 | 1.04 ± 0.21 | Treatment | 0.41 | 0.668 |
| Bread + gold KF | 0.80 ± 0.30 | 1.01 ± 0.10 | 0.97 ± 0.07 | 1.15 ± 0.21 | Brain region | 0.53 | 0.669 | |
| Bread + green KF | 1.34 ± 0.30 | 0.88 ± 0.10 | 0.88 ± 0.07 | 0.82 ± 0.21 | Treatment × regions | 1.73 | 0.181 | |
| SOD2 (relative expression) | Bread only | 0.96 ± 0.11 | 1.51 ± 0.15 | 1.00 ± 0.10 | 0.92 ± 0.28 | Treatment | 0.31 | 0.736 |
| Bread + gold KF | 1.09 ± 0.11 | 1.12 ± 0.15 | 0.87 ± 0.10 | 0.99 ± 0.28 | Brain region | 6.24 | 0.006 | |
| Bread + green KF | 1.11 ± 0.11 | 1.10 ± 0.15 | 0.92 ± 0.10 | 1.33 ± 0.28 | Treatment × regions | 0.91 | 0.516 | |
| AChE activity (nM H2O2/µg protein/min) | Bread only | 45.97 ± 1.02 | 93.19 ± 1.01 | 39.86 ± 1.02 | 28.48 ± 1.04 | Treatment | 2.90 | 0.086 |
| Bread + gold KF | 45.45 ± 1.02 | 93.20 ± 1.01 | 37.43 ± 1.02 | 24.96 ± 1.04 | Brain region | 3045.85 | <0.001 | |
| Bread + green KF | 46.63 ± 1.02 | 93.55 ± 1.01 | 38.02 ± 1.02 | 26.02 ± 1.04 | Treatment × regions | 1.77 | 0.172 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanon, A.P.; Giezenaar, C.; Roy, N.C.; Jayawardana, I.A.; Lomiwes, D.; Montoya, C.A.; McNabb, W.C.; Henare, S.J. Effects of Green and Gold Kiwifruit Varieties on Antioxidant Neuroprotective Potential in Pigs as a Model for Human Adults. Nutrients 2024, 16, 1097. https://doi.org/10.3390/nu16081097
Kanon AP, Giezenaar C, Roy NC, Jayawardana IA, Lomiwes D, Montoya CA, McNabb WC, Henare SJ. Effects of Green and Gold Kiwifruit Varieties on Antioxidant Neuroprotective Potential in Pigs as a Model for Human Adults. Nutrients. 2024; 16(8):1097. https://doi.org/10.3390/nu16081097
Chicago/Turabian StyleKanon, Alexander P., Caroline Giezenaar, Nicole C. Roy, Isuri A. Jayawardana, Dominic Lomiwes, Carlos A. Montoya, Warren C. McNabb, and Sharon J. Henare. 2024. "Effects of Green and Gold Kiwifruit Varieties on Antioxidant Neuroprotective Potential in Pigs as a Model for Human Adults" Nutrients 16, no. 8: 1097. https://doi.org/10.3390/nu16081097
APA StyleKanon, A. P., Giezenaar, C., Roy, N. C., Jayawardana, I. A., Lomiwes, D., Montoya, C. A., McNabb, W. C., & Henare, S. J. (2024). Effects of Green and Gold Kiwifruit Varieties on Antioxidant Neuroprotective Potential in Pigs as a Model for Human Adults. Nutrients, 16(8), 1097. https://doi.org/10.3390/nu16081097