The Ex Situ Conservation and Potential Usage of Crop Wild Relatives in Poland on the Example of Grasses




Abstract
:1. Introduction
2. Materials and Methods
- ✓
- ✓
- ✓
- recognizing (direct use) and potential usage of species in Poland were classified according to the following categories:
- ✓
- Fodder (FO)—plants which could be eaten by wild animals of farm livestock;
- ✓
- edible (ED)—plants used by humans as an additive in other dietary elements;
- ✓
- the biomass grasses (BG)—taxa can be used as sources of renewable energy (direct combustion, biogas, conversion to ethanol, etc.) or for different industrial purposes, i.e., pulp for paper production, as a component of construction or insulation elements;
- ✓
- amenity grasses (AG)—the possible (or actual) application is for home, sport, or other natural lawns, green surfaces, for landscape areas, road banks, etc. Grasses can be used as a specimen plant in perennial flower beds or in large groupings and mass plantings in gardens, parks, and recreation areas. Whole plants or panicles can be used in floristry in dry flower arrangements;
- ✓
- ecology (EC)—i.e., plants suitable to be grown in a wide range of soil conditions including waste and polluted areas, for land reclamation and/or habitat restoration and soil stabilization. Species for increasing biodiversity on arable lands, i.e., green areas as grassy strips on the fields (field margins).
- ✓
- Presence in long-term storage in the National Centre for Plant Genetic Resources in Poland and in the Kostrzyca Forest Gene Bank, as well as in other regional seed banks;
- ✓
- presence in botanical gardens and arboreta as listed in three studies: “Index Plantarum Polskich Kolekcji Dendrologicznych” (=Index Plantarum of the Polish dendrological collections) [40], “Index Plantarum of Outdoors Cultivated Herbaceous Plants in Poland” [41], and “Kolekcje Roślin Chronionych i Zagrożonych oraz Gatunków Objętych Konwencją Berneńską w Polskich Ogrodach Botanicznych” (=Collections of protected law and endangered plants and of species protected by the Bern Convention in the Polish Botanical Gardens) [42].
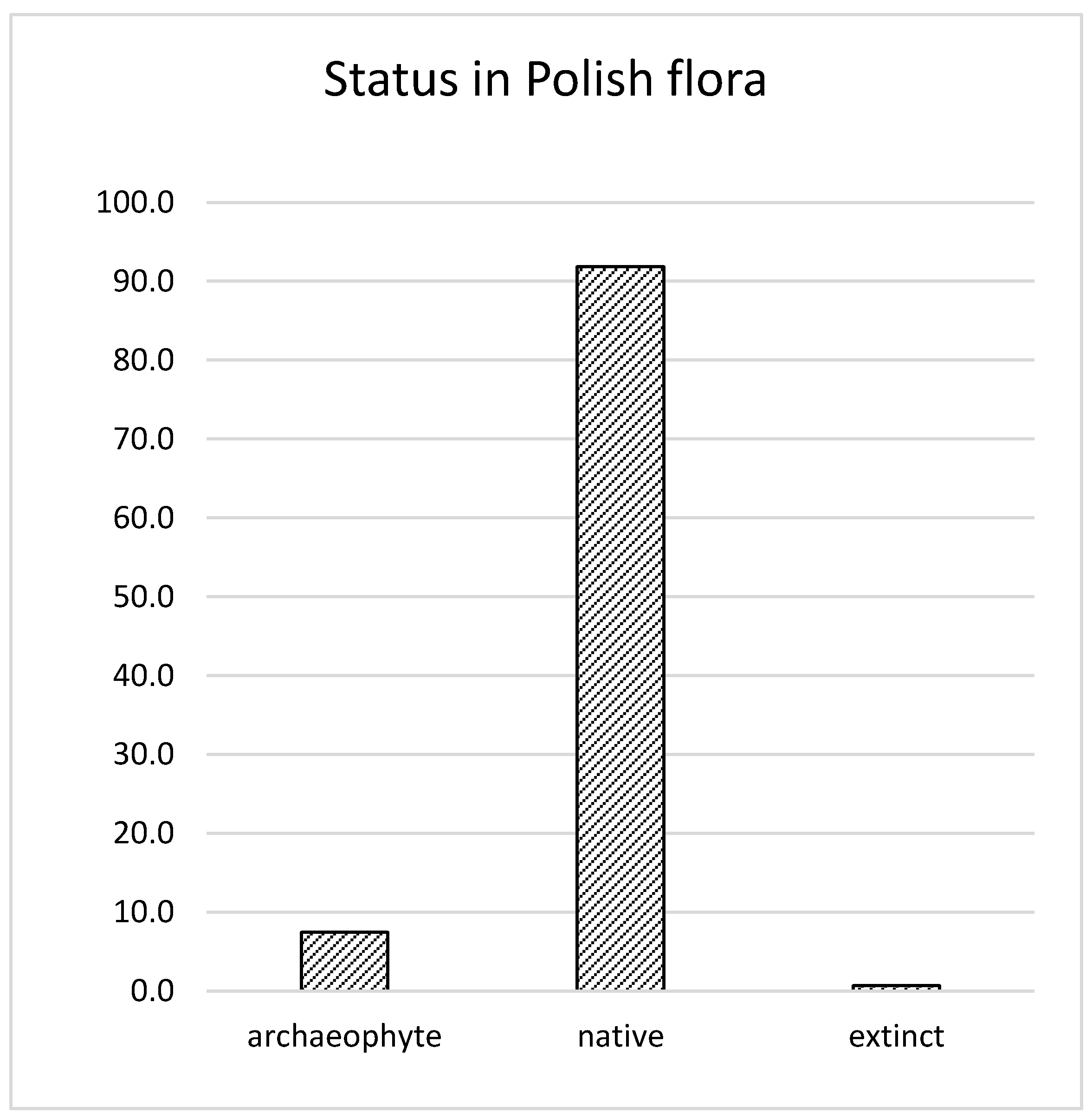
3. Results and Discussion
3.1. CWR in the Genetic Improvement of Other Species
3.2. The Potential of Crop Wild Relatives Direct Uses
3.2.1. Fodder and Edible Plants
3.2.2. Biomass Grasses
3.2.3. Amenity Grasses
3.2.4. Ecology
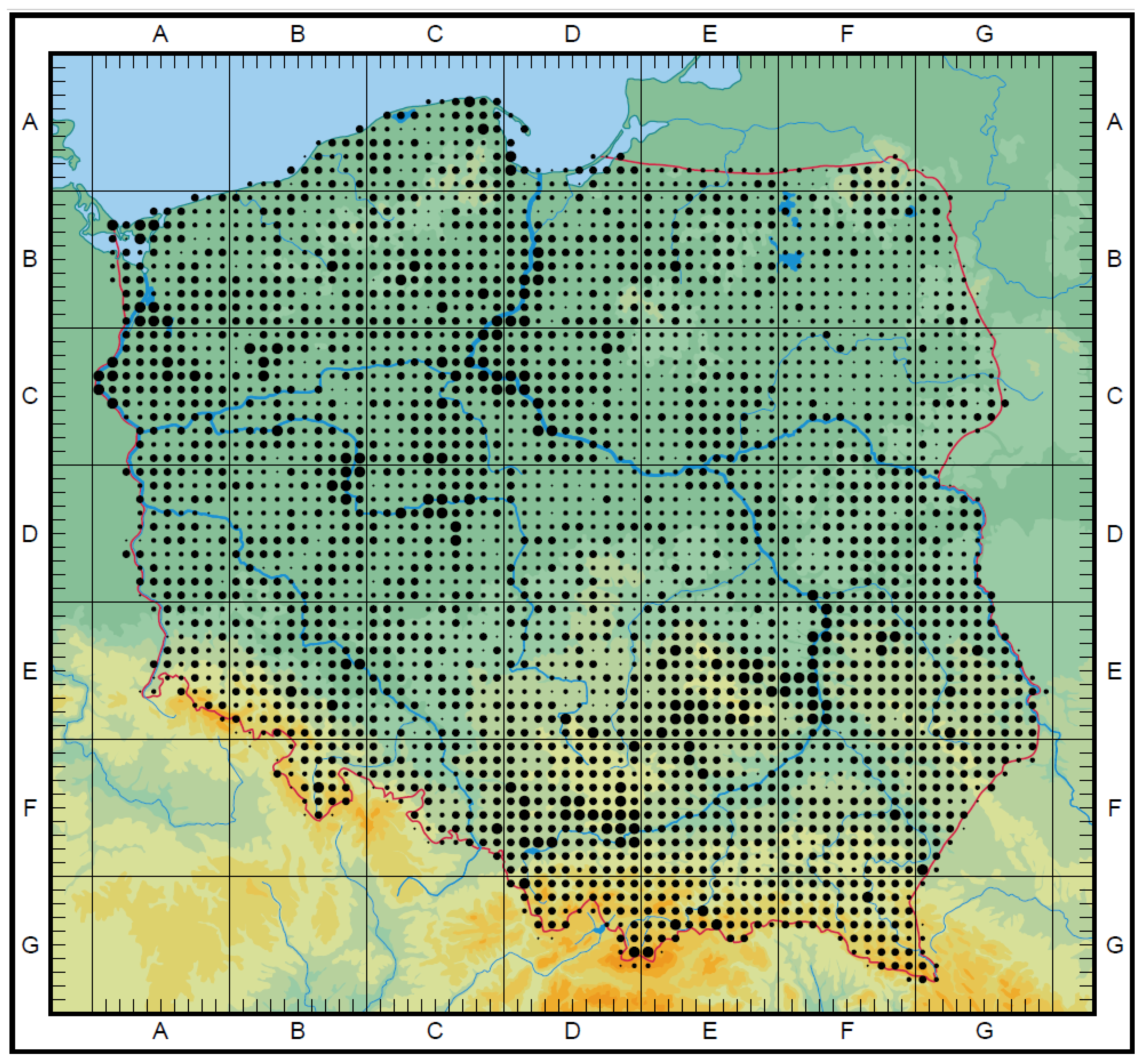
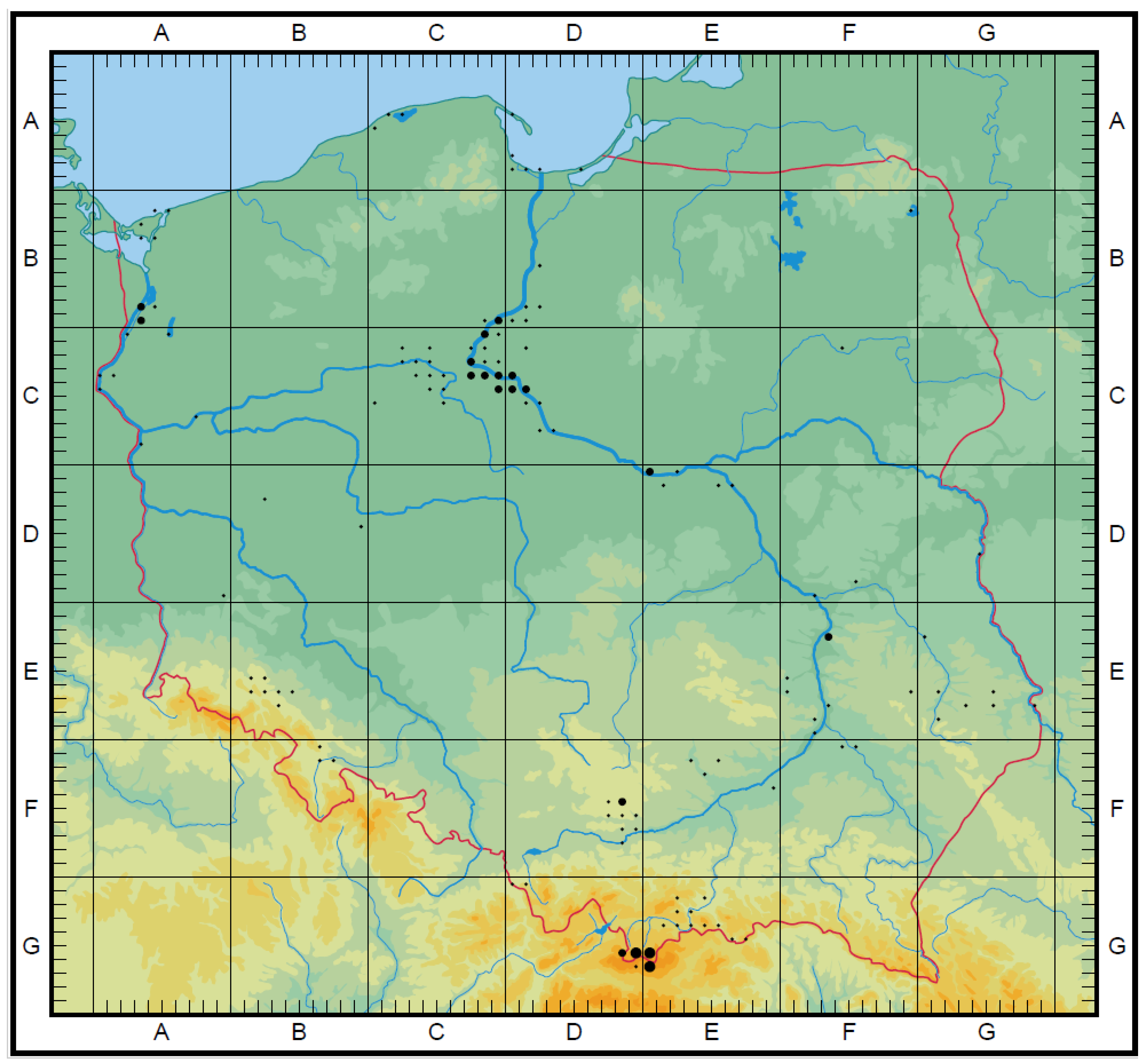
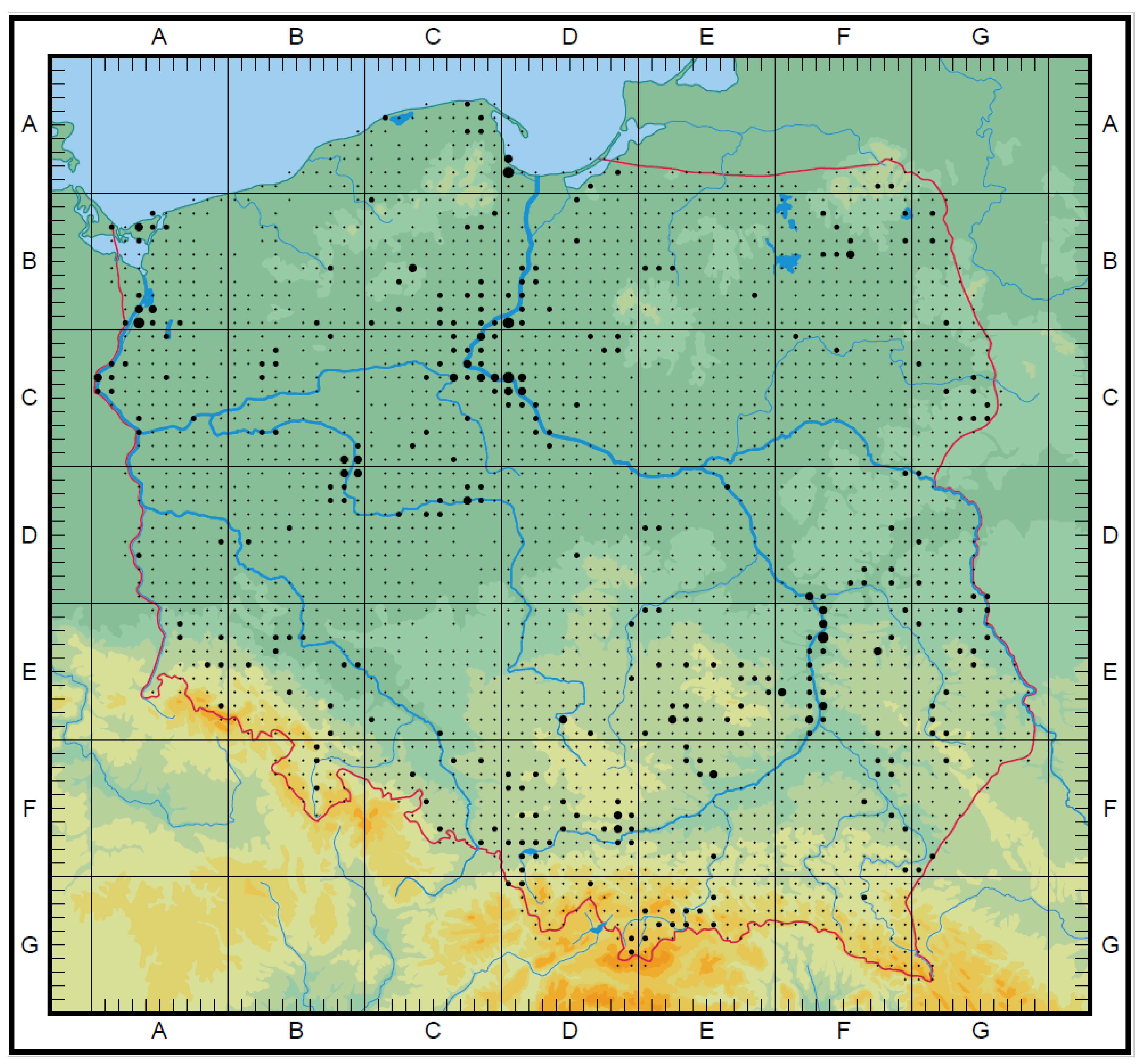
3.3. The Ex Situ Conservation in Gene Banks and Botanical Gardens
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
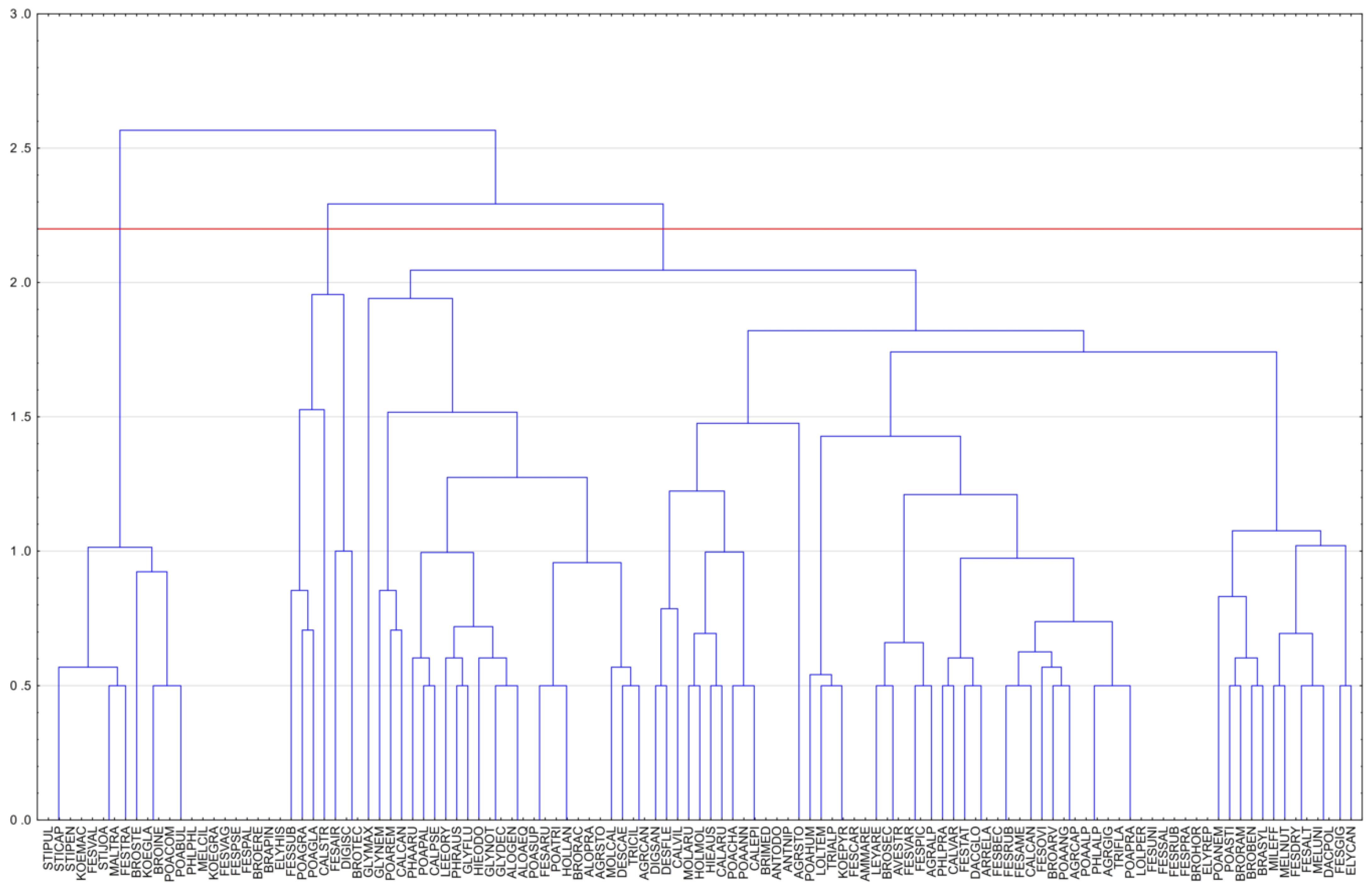{kind=link}
{kind=link}
| Taxon Name | Taxa Acronym | Floristic Status in Polish Flora | Potential Direct Use | Preferred Habitats | European Red List (Vascular Plants/Medicinal Plants) | Polish Red List/Polish Red Data Book | Ellenberg’s Indicator Values | Presence in Collections | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fodder | Edible | Amenity Grasses | Biomass, Fuels and Raw Material | Ecology | Light Value (L) | Soil Moisture Value (M) | Soil Acidity (pH) Value (R) | Botanical Gardens and Arboretum | Presence in Gene Banks | Seed Banks | |||||||
| NCPGR | KFGB | ||||||||||||||||
| Agrostis alpina Scop. | AGRALP | N | ● | 10, 11 | VU/VU | 5 | 3–4 | 3–5 | |||||||||
| Agrostis canina L. | AGRCAN | N | ● | ● | 4 | LC/- | 4 | 4 | 3 | ● | ● | ||||||
| Agrostis capillaris L. | AGRCAP | N | ● | 10 | 4 | 2–3 | 3–4 | ● | ● | ● | |||||||
| Agrostis gigantea Roth | AGRGIG | N | ● | ● | 2 | 4 | 3 | 3–4 | ● | ● | |||||||
| Agrostis rupestris All. | AGRRUP | N | ● | 10 | ● | ● | |||||||||||
| Agrostis stolonifera L. | AGRSTO | N | ● | ● | 2 | LC/- | 4 | 4 | 3–5 | ● | ● | ||||||
| Agrostis vinealis Schreb. | AGRSTO | N | ● | 3 | DD/- | 3 | 2 | 2–4 | ● | ||||||||
| Alopecurus aequalis Sobol. | ALOAEQ | N | ● | 2 | LC/- | 4 | 5 | 3–4 | ● | ||||||||
| Alopecurus arundinaceus Poir. In Lam. | ALOARU | N | ● | ● | 2, 10, 11, 12 | ● | |||||||||||
| Alopecurus geniculatus L. | ALOGEN | N | ● | 2 | LC/- | 4 | 5 | 3–4 | ● | ||||||||
| Alopecurus pratensis L. | ALOPRA | N | ● | ● | ● | 2 | LC/- | 4 | 4 | 4 | ● | ● | |||||
| Ammophila arenaria (L.) Link | AMMARE | N | ● | ● | ● | 13 | 5 | 3 | 5 | ● | ● | ||||||
| Anthoxanthum odoratum L. | ANTODO | N | ● | ● | 3, 4, 5, 6 | 4 | 3 | 3 | ● | ● | |||||||
| Anthoxanthum odoratum subsp. nipponicum (Honda) Tzvelev | ANTNIP | N | ● | ● | ● | 10, 11 | 4 | 3 | 3 | ||||||||
| Arrhenatherum elatius (L.) P. Beauv. ex J. Presl & C. Presl | ARRELA | N | ● | ● | ● | ● | ● | 2, 3 | LC/- | 4 | 3 | 4–5 | ● | ● | |||
| Avena strigosa Schreb. | AVESTR | A | ● | ● | ● | 4 | DD/- | 5 | 3 | 3–4 | ● | ● | |||||
| Beckmannia eruciformis (L.) Host | BECERU | A | ● | ● | ● | 2 | |||||||||||
| Brachypodium pinnatum (L.) P. Beauv. | BRAPIN | N | ● | ● | 1, 6 | 5 | 2 | 5 | ● | ● | |||||||
| Brachypodium sylvaticum (Huds.) P. Beauv. | BRASYL | N | ● | 5, 6 | 3 | 3–4 | 4–5 | ● | ● | ||||||||
| Briza media L. | BRIMED | N | ● | ● | 2 | 4 | 3 | 2–4 | ● | ● | |||||||
| Bromus arvensis L. | BROARV | A | ● | ● | 4 | VU/- | 4–5 | 2–3 | 3–4 | ● | |||||||
| Bromus benekenii (Lange) Trimen | BROBEN | N | ● | ● | 5, 6 | 3 | 3–4 | 5 | ● | ● | |||||||
| Bromus commutatus Schrad. | BROCUM | N | ● | ● | |||||||||||||
| Bromus erectus Huds. | BROERE | N | ● | ● | ● | 1 | 5 | 2 | 5 | ● | ● | ||||||
| Bromus hordeaceus L. | BROHOR | N | ● | 2 | 4 | 3 | 4 | ● | ● | ||||||||
| Bromus inermis Leyss. | BROINE | N | ● | ● | ● | ● | 1 | 5 | 2 | 4–5 | ● | ● | |||||
| Bromus racemosus L. | BRORAC | N | ● | ● | 1 | NT/- | 4 | 4 | 4 | ||||||||
| Bromus ramosus Huds. | BRORAM | N | ● | 5, 6 | VU/- | 3 | 3 | 5 | ● | ● | |||||||
| Bromus secalinus L. | BROSEC | A | ● | ● | 4 | 5 | 3 | 3–4 | ● | ● | ● | ||||||
| Bromus sterilis L. | BROSTE | A | ● | 4 | 5 | 2 | 4 | ● | ● | ||||||||
| Bromus tectorum L. | BROTEC | A | ● | ● | 4 | 5 | 2 | 3 | ● | ● | |||||||
| Calamagrostis arundinacea (L.) ROTH | CALARU | N | ● | ● | 5, 6, 7, 8 | 3 | 3 | 2–3 | ● | ● | |||||||
| Calamagrostis canescens (Weber) Roth | CALCAN | N | ● | ● | 2, 5 | 3 | 5 | 4 | ● | ||||||||
| Calamagrostis epigejos (L.) Roth | CALEPI | N | ● | 4 | 4 | 3 | 3 | ● | ● | ||||||||
| Calamagrostis pseudophragmites (Haller F.) Koeler | CALPSE | N | ● | ● | 2 | 4 | 4–5 | 5 | ● | ||||||||
| Calamagrostis stricta (Timm) Koeler | CALSTR | N | ● | ● | 2 | NT/- | 5 | 5 | 2–3 | ● | |||||||
| Calamagrostis varia (Schrad.) Host | CALVAR | N | ● | ● | 8, 11 | 4 | 2–3 | 5 | ● | ||||||||
| Calamagrostis villosa (Chaix) J. F. Gmel. | CALVIL | N | ● | ● | 5 | 4–3 | 3–4 | 1–2 | ● | ||||||||
| Corynephorus canescens (L.) P. Beauv. | CALCAN | N | ● | 9, 13 | 4 | 2 | 3–5 | ● | ● | ||||||||
| Dactylis glomerata L. * | DACGLO | N | ● | ● | ● | 2,3 | 4 | 3 | 4–5 | ● | ● | ● | |||||
| Dactylis polygama Horv. * | DACPOL | N | ● | ● | 5, 6 | 3 | 3 | 4 | ● | ● | |||||||
| Deschampsia caespitosa (L.) P. Beauv. | DESCAE | N | ● | ● | ● | ● | 2 | 3–5 | 4 | 3–4 | ● | ● | |||||
| Deschampsia flexuosa (L.) Trin. | DESFLE | N | ● | 5, 6, 7, 8 | 3–4 | 3 | 1–3 | ● | ● | ||||||||
| Deschampsia setacea (Huds.) Hack. | DESSET | E | ● | 2 | RE/EX | ||||||||||||
| Digitaria ischaemum (Schreb.) H. L. Mühl. | DIGISC | A | ● | 4 | 5 | 2 | 3 | ● | ● | ||||||||
| Digitaria sanguinalis (L.) Scop. | DIGSAN | A | ● | 4 | 4 | 3 | 2 | ● | ● | ||||||||
| Elymus caninus (L.) L. | ELYCAN | N | ● | 4 | LC/- | 3 | 4 | 4 | ● | ||||||||
| Elymus farctus (Viv.) Runemark ex Melderis * | ELYFAR | N | ● | 13 | -/CR | ● | ● | ● | |||||||||
| Elymus hispidus (Opiz) Melderis | ELYHIS | N | ● | 4 | 5 | 2 | 5 | ● | ● | ||||||||
| Elymus repens (L.) Gould | ELYREP | N | ● | ● | 4 | 4 | 3 | 3–5 | ● | ● | |||||||
| Festuca airoides Lam. | FESAIR | N | ● | 10 | 5 | 2 | 2 | ● | |||||||||
| Festuca altissima All. | FESALT | N | ● | ● | ● | 5, 6 | 3 | 3 | 3–4 | ● | ● | ||||||
| Festuca amethystina L. | FESAME | N | ● | ● | 10, 11 | EN/EN | 4 | 2–3 | 4 | ● | |||||||
| Festuca amethystina ssp. ritschlii * | FESRIT | N | ● | ● | 5, 6 | ||||||||||||
| Festuca arundinacea Schreb. | FESARU | N | ● | ● | ● | ● | 2, 12 | 4 | 3–4 | 4 | ● | ● | |||||
| Festuca beckeri (Hack.) Trautv. | FESBEC | N | ● | 3 | NT/- | 4 | 2–3 | 5 | ● | ● | |||||||
| Festuca carpathica F.Dietr. | FESCAR | N | ● | 11 | 5 | 3 | 5 | ● | |||||||||
| Festuca diffusa Dumort. | FESDIF | N | ● | 2 | |||||||||||||
| Festuca drymeia Mert. & W. D. J. Koch | FESDRY | N | ● | ● | 2 | 3 | 3 | 3–4 | |||||||||
| Festuca duriuscula L. Festuca rubra L. | FESRUB | N | ● | ● | ● | 2,3 | 4 | 2–4 | 5–6 | ||||||||
| Festuca duvalii (St. Yves) Stohr | FESDUV | N | ● | 3 | DD/- | ||||||||||||
| Festuca filiformis Pourr. | FESFIL | N | ● | ● | 2 | DD/- | 4 | 3–4 | ● | ● | |||||||
| Festuca gigantea (L.) Vill. | FESGIG | N | ● | 5 | 2–3 | 4 | 4 | ● | ● | ||||||||
| Festuca heterophylla Lam. | FESHET | N | ● | ● | 6 | LC/- | NT/- | ● | ● | ||||||||
| Festuca macutrensis Zapał. | FESMAC | N | ● | 1 | EN/EN | 2–3 | 4 | 5–6 | |||||||||
| Festuca nigrescens Lam. | FESNIG | N | ● | ● | ● | 2 | DD/ | ● | ● | ||||||||
| Festuca ovina L. * | FESOVI | N | ● | ● | ● | 3 | LC/- | 4 | 2 | 3–4 | ● | ● | |||||
| Festuca ovina L. var. vulgaris W.D.J.Koch subvar. guestphalica Hack. * | FESGUE | N | ● | ● | 3 | DD/- | 4 | 2 | 5–6 | ||||||||
| Festuca pallens Host | FESPAL | N | ● | 11 | 5 | 2 | 5 | ● | ● | ||||||||
| Festuca picta Kit. * | FESPIC | N | ● | 11 | 5 | 3 | 4 | ● | |||||||||
| Festuca pratensis Huds. | FESPRA | N | ● | ● | ● | 2, 3 | 4 | 3 | 4 | ● | ● | ||||||
| Festuca psammophila (Hack. ex Celak.) Fritsch | FESPSA | N | ● | ● | 3 | NT/- | 5 | 2 | ● | ||||||||
| Festuca pseudodalmatica Krajina ex Domin | FESDAL | N | ● | ● | 1 | CR/CR | ● | ||||||||||
| Festuca pseudovina Hack. ex Wiesb. | FESPSE | N | ● | ● | 3 | CR/CR | 5 | 2 | 5 | ● | |||||||
| Festuca rubra L. s. str. * | FESRUB | N | ● | ● | ● | 2 | LC/- | 4 | 2–4 | 4–6 | ● | ● | |||||
| Festuca rupicola Heuff. | FESRUP | N | ● | ● | 1 | 5 | 5 | ● | |||||||||
| Festuca salina Natho & Stohr * | FESSAL | N | ● | 12 | 4 | 2–4 | 4 | ||||||||||
| Festuca tatrae (Czakó) Degen | FESTAT | N | ● | ● | 10, 11 | 4 | 3 | 5 | ● | ● | |||||||
| Festuca trachyphylla (Hack.) Krajina | FESTRA | N | ● | ● | 1 | 5 | 1–2 | 5–6 | ● | ||||||||
| Festuca unifaria Dumort. * | FESUNI | N | ● | ● | ● | 2 | 4 | 2–4 | 4 | ||||||||
| Festuca vaginata Waldst. & Kit. ex Willd. | FESVAG | N | ● | ● | 2 | DD/- | 5 | 2 | 5 | ● | ● | ||||||
| Festuca valesiaca Schleich. ex Gaudin | FESVAL | N | ● | 1 | VU/- | 5 | 1 | 5 | ● | ● | |||||||
| Festuca varia Haenke | FESVAR | N | ● | ● | 11 | 5 | 3 | 4 | ● | ||||||||
| Festuca villosa Schweigg. | FESVIL | N | ● | ● | |||||||||||||
| Glyceria declinata Bréb. | GLYDEC | N | ● | 2 | LC/- | 4 | 5 | 4 | |||||||||
| Glyceria fluitans (L.) R. Br. | GLYFLU | N | ● | 2 | LC/- | 4 | 6–5 | 4 | ● | ● | |||||||
| Glyceria lithuanica (Gorski) Gorski | GLYLIT | N | ● | 7 | CR/CR | ● | |||||||||||
| Glyceria maxima (Hartm.) Holmb. | GLYMAX | N | ● | ● | ● | 2 | LC/- | 4 | 6 | 5 | ● | ● | |||||
| Glyceria nemoralis (R. Uechtr.) R. Uechtr. & Körn. | GLYNEM | N | ● | 6 | LC/- | 3 | 5 | 5 | |||||||||
| Glyceria notata Chevall. | GLYNOT | N | ● | 2 | 4 | 5 | 4 | ● | |||||||||
| Glyceria striata (Lam.) Hitchc. | GLYSTR | A | ● | 2 | |||||||||||||
| Hierochloë australis (Schrad.) Roem. & Schult. | HIEAUS | N | ● | ● | ● | 5, 6, 7, 8 | VU/- | 3 | 3 | 3 | ● | ● | |||||
| Hierochloë odorata (L.)P. Beauv. | HIEODO | N | ● | 1,3, 5, 6 | VU/- | 4 | 4–5 | 4 | ● | ● | |||||||
| Holcus lanatus L. | HOLLAN | N | ● | 2 | 4 | 4 | 4 | ● | ● | ||||||||
| Holcus mollis L. | HOLMOL | N | ● | 9 | 3–4 | 3–4 | 3 | ● | |||||||||
| Koeleria glauca(Spreng.) Dc. | KOEGLA | N | ● | 3 | 5 | 2 | 4–5 | ● | ● | ||||||||
| Koeleria grandis Besser ex Gorski * | KOEGRA | N | ● | 1 | DD/- | 5 | 2 | 5 | ● | ● | |||||||
| Koeleria macrantha (Ledeb.) Schult. | KOEMAC | N | ● | ● | 3 | 5 | 1 | 5 | ● | ● | ● | ||||||
| Koeleria pyramidata (Lam.) P. Beauv. * | KOEPYR | N | ● | ● | 1 | VU/- | 5 | 3 | 5 | ● | ● | ||||||
| Leersia oryzoides (L.) Sw. | LEEORY | N | ● | 2 | LC/- | NT/- | 5–4 | 5 | 4 | ● | |||||||
| Leymus arenarius (L.) Hochst. * | LEYARE | N | ● | ● | ● | ● | 13 | 5 | 2–3 | 3–4 | ● | ● | |||||
| Lolium perenne L. | LOLPER | N | ● | ● | ● | 4 | LC/- | 4 | 3 | 4 | ● | ● | |||||
| Lolium remotum Schrank | LOLREM | A | ● | 4 | CR/- | ● | ● | ||||||||||
| Lolium temulentum L. | LOLTEM | A | ● | 4 | LC/- | VU/- | 5 | 3 | 4–5 | ● | ● | ||||||
| Melica ciliata L. | MELCIL | N | ● | 1 | 5 | 2 | 5 | ● | ● | ● | |||||||
| Melica nutans L. | MELNUT | N | ● | ● | ● | 5, 6 | 2–3 | 3 | 4 | ● | ● | ||||||
| Melica picta K. Koch | MELPIC | N | ● | ● | 5, 6 | CR/CR | ● | ||||||||||
| Melica transsilvanica Schur | MALTRA | N | ● | 1 | NT/- | 5 | 1 | 4–5 | ● | ● | ● | ||||||
| Melica uniflora Retz. | MELUNI | N | ● | ● | 5, 6 | 3 | 3 | 4 | ● | ||||||||
| Milium effusum L. | MILEFF | N | ● | ● | 5, 6 | 2–3 | 3–4 | 4 | ● | ● | |||||||
| Molinia arundinacea Schrank * | MOLARU | N | ● | ● | 2 | 3 | 3–4 | 3 | ● | ● | |||||||
| Molinia caerulea (L.) Moench * | MOLCAL | N | ● | ● | 2 | 4 | 4–5 | 1–5 | ● | ● | |||||||
| Phalaris arundinacea L. | PHAARU | N | ● | ● | ● | ● | ● | 2 | LC/- | 4 | 5 | 4–5 | ● | ● | |||
| Phleum alpinum L. | PHLALP | N | ● | 10, 11 | 4 | 3 | 3–4 | ● | ● | ||||||||
| Phleum bertolonii Dc. * (P. hubbardii D. Kovats) | PHLBER | N | ● | ● | ● | 3 | 4 | 2 | 5–6 | ||||||||
| Phleum hirsutum Honck. | PHLHIR | N | ● | ● | 10, 11 | ||||||||||||
| Phleum phleoides (L.) H. Karst. | PHLPHL | N | ● | 2 | 5 | 2 | 5 | ● | ● | ● | |||||||
| Phleum pratense L. | PHLPRA | N | ● | ● | ● | 2, 3 | LC/- | 4 | 2–3 | 4–5 | ● | ● | |||||
| Phragmites australis (Cav.) Trin. ex Steud. | PHRAUS | N | ● | ● | ● | ● | 12 | LC/- | 4–5 | 5–6 | 4 | ● | |||||
| Poa alpina L. | POAALP | N | ● | 10, 11 | LC/- | 4 | 3 | 3–4 | ● | ● | |||||||
| Poa angustifolia L. | POAANG | N | ● | 10, 11 | 4 | 2–3 | 4–5 | ● | |||||||||
| Poa annua L. | POAANN | N | ● | 4 | 5–3 | 3 | 4 | ● | |||||||||
| Poa bulbosa L. | POABUL | N | ● | 1 | NT/- | 5 | 2 | 5 | ● | ● | |||||||
| Poa chaixii Vill. | POACHA | N | ● | 10, 11 | 4 | 3 | 2–3 | ● | ● | ||||||||
| Poa compressa L. | POACOM | N | ● | ● | ● | 1 | 5 | 2 | 5 | ● | ● | ||||||
| Poa glauca Vahl | POAGLA | N | ● | ● | 11 | 5 | 3–4 | 3 | ● | ● | |||||||
| Poa granitica Braun–Blanq. | POAGRA | N | ● | 10 | DD/- | NT/NT | 5 | 4 | 2–3 | ||||||||
| Poa humilis Ehrh. ex Hoffm. | POAHUM | N | ● | 1, 2, 13 | 5 | 4–3 | 5 | ||||||||||
| Poa laxa Haenke | FESSUB | N | ● | ● | 10 | 5 | 3 | 2–3 | ● | ||||||||
| Poa molinerii Balb. | POAMOL | N | ● | 10, 11 | 5 | 3 | 2–3 | ||||||||||
| Poa nemoralis L. | POANEM | N | ● | ● | 5, 6 | 3 | 2–3 | 4–5 | ● | ● | |||||||
| Poa nobilis Skalińska | PONNOB | N | ● | 10 | DD/DD | ||||||||||||
| Poa palustris L. | POAPAL | N | ● | ● | 2 | 4 | 4–5 | 4–5 | ● | ● | ● | ||||||
| Poa pratensis L. | POAPRA | N | ● | ● | 2 | LC/- | 4 | 3 | 4 | ● | ● | ||||||
| Poa remota Forselles | POAREM | N | ● | 10, 11 | 3 | 4–5 | 4–5 | ||||||||||
| Poa stiriaca Fritsch & Hayek | POASTI | N | ● | 6 | VU/VU | 3 | 3 | 4–5 | ● | ● | |||||||
| Poa supina Schrad. | POASUP | N | ● | ● | 10 | 4 | 3–4 | 4 | ● | ||||||||
| Poa trivialis L. | POATRI | N | ● | 2 | 4 | 4 | 4 | ● | ● | ||||||||
| Stipa capillata L. | STICAP | N | ● | ● | 1 | VU/- | 5 | 1 | 5 | ● | ● | ● | |||||
| Stipa eriocaulis Borbás * | STIEUR | N | ● | 1 | RE/- | ● | |||||||||||
| Stipa joannis Čelak. s. s. | STIJOA | N | ● | ● | 1 | VU/VU | 5 | 1 | 5–4 | ● | ● | ● | |||||
| Stipa pennata L. (Stipa borysthenica Klokov) * | STIPEN | N | ● | ● | 1 | CR/CR | 5 | 1 | 5 | ● | ● | ||||||
| Stipa pulcherrima K. Koch | STIPUL | N | ● | ● | 1 | VU/VU | 5 | 1 | 5 | ● | ● | ● | |||||
| Trisetum alpestre (Host) P. Beauv. | TRIALP | N | ● | 10, 11 | 5 | 3 | 5 | ● | |||||||||
| Trisetum ciliare (Kit.) Domin | TRICIL | N | ● | 10, 11 | 4 | 4 | 3 | ||||||||||
| Trisetum flavescens (L.) P. Beauv. | TRIFLA | N | ● | ● | 11 | 4 | 3 | 4 | ● | ● | |||||||
| Trisetum sibiricum Rupr. | TRISIB | N | ● | 2 | NT/NT | 4 | 4 | ||||||||||
| Total | 116 | 82 | 10 | 8 | |||||||||||||
References
- Vavilov, N.I. The law of homologous series in variation. J. Genet. 1922, 12, 47–89. [Google Scholar] [CrossRef]
- Harlan, J.R.; de Wet, J.M. Toward a rational classification of cultivated plants. Taxon 1971, 20, 509–517. [Google Scholar] [CrossRef]
- Maxted, N.; Ford-Lloyd, B.V.; Jury, S.; Kell, S.; Scholten, M. Towards a definition of a crop wild relative. Biodivers. Conserv. 2006, 15, 2673–2685. [Google Scholar] [CrossRef]
- Maxted, N.; Avagyan, A.; Frese, L.; Iriondo, J.M.; Magos Brehm, J.; Singer, A.; Kell, S.P. ECPGR Concept for In Situ Conservation of Crop Wild Relatives in Europe; Wild Species Conservation in Genetic Reserves Working Group, European Cooperative Programme for Plant Genetic Resources: Rome, Italy, 2015; Available online: http://www.pgrsecure.org/documents/Concept_v2.pdf (accessed on 10 December 2020).
- Fielder, H.; Brotherton, P.; Hosking, J.; Hopkins, J.J.; Ford-Lloyd, B.; Maxted, N. Enhancing the Conservation of Crop Wild Relatives in England. PLoS ONE 2015, 10, e0130804. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, H. The National Crop Wild Relative Strategy Report for Finland; MTT: Jokioinen, Finland, 2013; p. 94. [Google Scholar]
- Landucci, F.; Panella, L.; Lucarini, D.; Gigante, D.; Donnini, D.; Kell, S.; Maxted, N.; Venanzoni, R.; Negri, V. A Prioritized Inventory of Crop Wild Relatives and Wild Harvested Plants of Italy. Crop Sci. 2014, 54, 1628–1644. [Google Scholar] [CrossRef]
- Magos Brehm, J.; Maxted, N.; Ford-Lloyd, B.V.; Martins-Louçao, M.A. National inventories of crop wild relatives and wild harvested plants: Case–Study for Portugal. Genet. Resour. Crop Evol. 2008, 55, 779–796. [Google Scholar] [CrossRef]
- Taylor, N.G.; Kell, S.P.; Holubec, V.; Parra-Quijano, M.; Chobot, K.; Maxted, N. A systematic conservation strategy for crop wild relatives in the Czech Republic. Divers. Distrib. 2017, 23, 448–462. [Google Scholar] [CrossRef]
- Dostatny, D.F.; Dajdok, Z. (Eds.) Dzikie Gatunki Pokrewne Roślinom Uprawnym Występujące w Polsce. Lista, Zasoby i Zagrożenia (Crop Wild Relatives Occurring in Poland. Checklist, Resources and Threats); Wydawnictwo Kontekst: Radzików-Poznań, Poland, 2020; p. 420. [Google Scholar]
- Teso, M.L.R.; Lamas, E.T.; Parra-Quijano, M.; de la Rosa, L.; Fajardo, J.; Iriondo, J.M. National inventory and prioritization of crop wild relatives in Spain. Genet. Resour. Crop Evol. 2018, 65, 1237–1253. [Google Scholar] [CrossRef]
- Contreras-Toledo, A.R.; Cortés-Cruz, M.A.; Costich, D.; de Lourdes Rico-Arce, M.; Brehm, J.M.; Maxted, N. A crop wild relative inventory for Mexico. Crop Sci. 2018, 58, 1292–1305. [Google Scholar] [CrossRef] [Green Version]
- Allen, E.; Gaisberger, H.; Brehm, J.M.; Maxted, N.; Thormann, I.; Lupupa, T.; Kell, S.P. A crop wild relative inventory for Southern Africa: A first step in linking conservation and use of valuable wild populations for enhancing food security. Plant Genet. Resour. 2019, 17, 128–139. [Google Scholar] [CrossRef]
- Mponya, N.K.; Magombo, Z.L.; Pungulani, L.; Brehm, J.M.; Maxted, N. Development of a prioritised checklist of crop wild relatives for conservation in Malawi. Afr. Crop Sci. J. 2020, 28, 279–311. [Google Scholar] [CrossRef]
- Kell, S.; Qin, H.; Chen, B.; Ford-Lloyd, B.; Wei, W.; Kang, D.; Maxted, N. China’s crop wild relatives: Diversity for agriculture and food security. Agric. Ecosyst. Environ. 2015, 209, 138–154. [Google Scholar] [CrossRef]
- Newton, A.C.; Johnson, S.N.; Gregory, P.J. Implications of climate change for diseases, crop yields and food security. Euphytica 2011, 179, 3–18. [Google Scholar] [CrossRef]
- Miraglia, M.; Marvin, H.J.P.; Kleter, G.A.; Battilani, P.; Brera, C.; Coni, E.; Filippi, L. Climate change and food safety: An emerging issue with special focus on Europe. Food Chem. Toxicol. 2009, 47, 1009–1021. [Google Scholar] [CrossRef]
- Tripathi, A.; Tripathi, D.K.; Chauhan, D.K.; Kumar, N.; Singh, G.S. Paradigms of climate change impacts on some major food sources of the world: A review on current knowledge and future prospects. Agric. Ecosyst. Environ. 2016, 216, 356–373. [Google Scholar] [CrossRef]
- Ford-Lloyd, B.V.; Schmidt, M.; Armstrong, S.J.; Barazani, O.; Engels, J.; Hadas, R.; Hammer, K.; Kell, S.; Kang, D.; Khoshbakht, K.; et al. Crop Wild Relatives—Undervalued, underutilized and under threat? BioScience 2011, 61, 559–565. [Google Scholar] [CrossRef] [Green Version]
- Maxted, N.; Magos Brehm, J.; Kell, S.P. Resource Book for the Preparation of National Plans for Conservation of Crop Wild Relatives and Landraces; FAO: Rome, Italy, 2013; p. 463. [Google Scholar]
- Jarvis, A.; Lane, A.; Hijmans, R.J. The effect of climate change on crop wild relatives. Agric. Ecosyst. Environ. 2008, 126, 13–23. [Google Scholar] [CrossRef]
- Redden, R.J.; Yadav, S.S.; Maxted, N.; Dulloo, M.E.; Guarino, L.; Smith, P. (Eds.) Crop Wild Relatives and Climate Change; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Phillips, J.; Brehm, J.M.; van Oort, B.; Asdal, Å.; Rasmussen, M.; Maxted, N. Climate change and national crop wild relative conservation planning. Ambio 2017, 46, 630–643. [Google Scholar] [CrossRef]
- Maxted, N.; Amri, A.; Castañeda-Álvarez, N.P.; Dias, S.; Dulloo, M.E.; Fielder, H.; Ford-Lloyd, B.V.; Iriondo, J.M.; Magos Brehm, J.; Nilsen, L.B.; et al. Joining up the Dots: A Systematic Perspective of Crop Wild Relative Conservation and Use. In Enhancing Crop Genepool Use: Capturing Wild Relative and Landrace Diversity for Crop Improvement; Maxted, N., Ehsan Dulloo, M., Ford-Lloyd, B., Eds.; CAB International: Wallingford, UK, 2016; pp. 87–124. [Google Scholar]
- Boller, B.; Schubiger, F.; Kölliker, R. Red Clover. In Fodder Crops and Amenity Grasses; Springer: New York, NY, USA, 2010; pp. 439–455. [Google Scholar]
- Chorley, G. The agricultural revolution in northern Europe, 1750–1880: Nitrogen, Legumes, and Crop Productivity. Econ. Hist. Rev. 1981, 34, 71–93. [Google Scholar]
- Drobnik, J. Manna polska od XVI do XIX w. pod względem botanicznym i leczniczym. Polish manna grass in the 16th to 19th centuries: A botanical and medicinal approach. Etnobiologia Pol. 2015, 5, 67–87. [Google Scholar]
- Hozyasz, K. From rediscovered “niche” to “mainstream”—Glyceria fluitans as a candidate grain for manufacturing premium food products. PlantsPeoplePlanet 2020, 2, 104–106. [Google Scholar] [CrossRef]
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Flowering Plants and Pteridophytes of Poland. A Checklist; Instytut Ochrony Przyrody, Polska Akademia Nauk: Kraków, Poland, 2002; p. 442. [Google Scholar]
- The Plant List. The Plant List—A Working List of all Plant Species. Available online: http://www.theplantlist.org (accessed on 10 December 2020).
- COBORU. 2020. Available online: https://coboru.gov.pl/ (accessed on 10 December 2020).
- Polish Nurserymen Association. Available online: http://zszp.pl/en/strona=glowna–en/ (accessed on 10 December 2020).
- Tokarska-Guzik, B.; Dajdok, Z.; Zając, M.; Zając, A.; Urbisz, A.; Danielewicz, W.; Hołdyński, C. Rośliny Obcego Pochodzenia w Polsce; Generalna Dyrekcja Ochrony Środowiska: Warszawa, Poland, 2012.
- Matuszkiewicz, W. Przewodnik do Oznaczania Zbiorowisk Roślinnych Polski; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2014; p. 540. [Google Scholar]
- Zarzycki, K.; Trzcińska–Tacik, H.; Różański, W.; Szeląg, Z.; Wołek, J.; Korzeniak, U. Ekologiczne Liczby Wskaźnikowe Roślin Naczyniowych Polski; Instytut Botaniki im. W. Szafera PAN: Kraków, Poland, 2002. [Google Scholar]
- Bilz, M.; Lansdown, R.; Kell, S.; Maxted, N. European Red List of Vascular Plants; Publications Office of the European Union: Luxembourg, 2011; p. 126. [Google Scholar]
- Allen, D.; Bilz, M.; Leaman, D.J.; Miller, R.M.; Timoshyna, A.; Window, J. European Red List of Medicinal Plants; Publications Office of the European Union: Luxembourg, 2014. [Google Scholar]
- Kaźmierczakowa, R.; Bloch-Orłowska, J.; Celka, Z.; Cwener, A.; Dajdok, Z.; Michalska-Hejduk, D.; Pawlikowski, P.; Szczęśniak, E.; Ziarnek, K. Polska Czerwona Lista Paprotników i Roślin Kwiatowych; Instytut Ochrony Przyrody Polskiej Akademii Nauk: Kraków, Poland, 2016; p. 48. [Google Scholar]
- Kaźmierczakowa, R.; Zarzycki, K.; Mirek, Z. Polska Czerwona Księga Roślin. Paprotniki i Rośliny Naczyniowe; Wyd. 3; Instytut Ochrony Przyrody, Polska Akademia Nauk: Kraków, Poland, 2014; p. 895. [Google Scholar]
- Nowak, T.; Baran, B.; Bojarczuk, T.; Dąbrowski, K.; Gawryś, W.; Grzeszczak-Nowak, H.; Jerzak, E.; Kałuża, A.; Kazimierska, K.; Kozłowska-Kalisz, J.; et al. Index Plantarum Polskich Kolekcji Dendrologicznych; Part 1; Prace Ogrodu Botanicznego Uniwersytetu Wrocawskiego: Wrocław, Poland, 1999; Volume 5, p. 306. [Google Scholar]
- Nowak, T.J.; Gawryś, W.; Marcinkowski, J. Index Plantarum of Outdoors Cultivated Herbaceous Plants in Poland. Pr. Ogrodu Bot. Uniw. Wrocławskiego 2000, 6, 5–399. [Google Scholar]
- Puchalski, J.; Gawryś, W. Kolekcje roślin chronionych i zagrożonych oraz gatunków objętych Konwencją Berneńską w polskich ogrodach botanicznych. Biul. Ogrodów Bot. Muzeów I Zbior. 2007, 16, 47–184. [Google Scholar]
- Zając, A.; Zając, M. Atlas Rozmieszczenia Roślin Naczyniowych w Polsce (Distribution Atlas of Vascular Plants in Poland); Pracownia Chorologii Komputerowej Instytutu Botaniki Uniwersytetu Jagiellońskiego: Kraków, Poland, 2001. [Google Scholar]
- Ellenberg, H.; Dull, R.; Wirth, V.; Paulißen, D.W. Zeigerwerte von Pflanzen in Mitteleuropa, 2nd ed.; Verlag Erich GoltzeKG: Goettingen, Germany, 1991; p. 248. [Google Scholar]
- Diekmann, M. Species indicator values as an important tool in applied plant ecology—A review. Basic Appl. Ecol. 2003, 4, 493–506. [Google Scholar] [CrossRef]
- Schaffers, A.; Sýkora, K. Reliability of Ellenberg indicator values for moisture, nitrogen and soil reaction: A comparison with field measurements. J. Veg. Sci. 2000, 11, 225–244. [Google Scholar] [CrossRef]
- Thompson, K.; Hodgson, J.G.; Grime, J.P.; Rorison, I.H.; Band, S.R.; Spencer, R.E. Ellenberg numbers revisited. Phytocoenologia 1993, 23, 277–289. [Google Scholar] [CrossRef]
- Ewald, J. The sensitivity of Ellenberg indicator values to the completeness of vegetation relevés. Basic Appl. Ecol. 2003, 4, 507–513. [Google Scholar] [CrossRef]
- Pawłowski, B. Remarques sur l’endémisme dans la flore des Alpes et des Carpates. Vegetatio 1970, 21, 181–243. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Araújo, M.B.; Jepson, P.; Ladle, R.J.; Watson, J.E.M.; Willis, K.J. Conservation biogeography: Assessment and prospect. Divers. Distrib. 2005, 11, 3–23. [Google Scholar] [CrossRef]
- Crisci, J.V.; Sala, O.E.; Katinas, L.; Posadas, P. Bridging historical and ecological approaches in biogeography. Aust. Syst. Bot. 2006, 19, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Schnittler, M.; Günther, K.F. Central European vascular plants requiring priority conservation measures–an analysis from national Red Lists and distribution maps. Biodivers. Conserv. 1999, 8, 891–925. [Google Scholar] [CrossRef]
- Galera, H.; Puchalski, J.; Gawryś, W. Polskie kolekcje roślin chronionych i zagrożonych oraz endemitów i reliktów. Część II. Taksony zagrożone, endemity i relikty. Biul. Ogrodów Bot. Muzeów I Zbior. 2000, 9, 19–41. [Google Scholar]
- Mirek, Z.; Piękoś-Mirkowa, H. Fitogeograficzne aspekty endemizmu w Polsce. Wiadomości Bot. 2009, 53, 7–30. [Google Scholar]
- Piękoś-Mirkowa, H.; Mirek, Z. Zagrożenie i ochrona endemicznych roślin naczyniowych występujących w Polsce. Chrońmy Przyr. Ojczystą 2010, 66, 15–26. [Google Scholar]
- Puchalski, J.; Niemczyk, M.; Walerowski, P.; Podyma, W.; Kapler, A. Seed banking of Polish endangered plants–the FlorNatur Project. Biodivers. Res. Conserv. 2014, 34, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Kiedrzyński, M.; Zielińska, K.M.; Kiedrzyńska, E.; Jakubowska-Gabara, J. Regional climate and geology affecting habitat availability for a relict plant in a plain landscape: The case of Festuca amethystina L. in Poland. Plant Ecol. Divers. 2015, 8, 331–341. [Google Scholar] [CrossRef]
- Łazarski, G. Festuca amethystina (Poaceae)–A species new for the Płaskowyż Jędrzejowski plateau. Fragm. Florist. Geobot. Pol. 2016, 23, 370–374. [Google Scholar]
- Klichowska, E.; Nobis, M. Stipa pennata subsp. ceynowae (Poaceae, Pooideae), a new taxon from Central Europe. PhytoKeys 2017, 83, 75. [Google Scholar] [CrossRef] [Green Version]
- Klichowska, E.; Ślipiko, M.; Nobis, M.; Szczecińska, M. Development and characterization of microsatellite markers for endangered species Stipa pennata (Poaceae) and their usefulness in intraspecific delimitation. Mol. Biol. Rep. 2018, 45, 639–643. [Google Scholar] [CrossRef] [Green Version]
- Rewicz, A.; Tomczyk, P.P.; Kiedrzyński, M.; Zielińska, K.M.; Jędrzejczyk, I.; Rewers, M.; Rewicz, T. Morphometric traits in the fine-leaved fescues depend on ploidy level: The case of Festuca amethystina L. PeerJ 2018, 6, e5576. [Google Scholar] [CrossRef] [Green Version]
- Szafer, W. Zarys Geografii Roślin; PWN: Warszawa, Poland, 1952. [Google Scholar]
- Podbielkowski, Z. Fitogeografia Części Świata; Wyd. Naukowe PWN: Warszawa, Poland, 2002; pp. 400 (I tom) + 236 (II tom). [Google Scholar]
- EU Directive 1992. Dyrektywa 1992. Dyrektywa Rady 92/43/EWG z dnia 21 maja 1992 r. o ochronie siedlisk przyrodniczych oraz dzikiej fauny i flory z późniejszymi zmianami. Available online: https://eur-lex.europa.eu/legal-content/PL/TXT/PDF/?uri=CELEX:01992L0043-20070101&from=EN (accessed on 10 December 2020).
- Czaplicki, A.; Plich, J.; Makowska, K.; Zimny, J. Androgeneza Jako Narzędzie do Poszerzania Puli Genetycznej Pszenicy Ozimej. 2016. Available online: http://pw.ihar.edu.pl/wp–content/uploads/2018/12/636e919349_Wojslawice.Czaplicki.PW.zad.2.1.–2016.pdf (accessed on 20 November 2020).
- Jakimur, N.S.; Snapp, S.S.; Murphy, K.; Jones, S.S. Agronomic assessment of perennial wheat and perennial rye as a cereal crop. Agron. J. 2012, 104, 1716–1726. [Google Scholar]
- Hayes, R.C.; Newell, M.T.; DeHaan, L.R.; Murphy, K.M.; Crane, S.; Norton, M.R.; Wade, L.J.; Newberry, M.; Fahim, M.; Jones, S.S.; et al. Perennial cereal crops: An initial evaluation of wheat derivatives. Field Crop. Res. 2012, 133, 68–89. [Google Scholar] [CrossRef]
- Verushkine, S.; Shechurdine, A. Hybrids between wheat and couch grass: Fertile Triticum–Agropyrum hybrids of great scientific and practical interest. J. Hered. 1933, 24, 329–335. [Google Scholar] [CrossRef]
- Cai, Y.; Xiang, F.; Zhi, D.; Liu, H.; Xia, G. Genotyping of somatic hybrids between Festuca arundinacea Schreb. and Triticum aestivum L. Plant Cell Rep. 2007, 26, 1809–1819. [Google Scholar] [CrossRef]
- Humphreys, M.W.; Zwierzykowski, Z. Festulolium, a century of research and breeding and its increased relevance in meeting the requirements for multifunctional grassland agriculture. Biol. Plant. 2020, 64, 578–590. [Google Scholar] [CrossRef]
- Łapiński, B.; Nita, Z.; Szołkowska, A.; Wieczorek, P. Mieszaniec owsa uprawnego z dzikim gatunkiem Avena macrostachya jako nowe źródło zmienności do poprawy jakości plonu owsa nagoziarnistego. Biul. Ihar 2013, 270, 43–54. [Google Scholar]
- Hanneberg, M.; Klawitter, M.; Kozłowski, J.; Marciniak, J.; Skrzydlewska, E. Zatrucia Roślinami Wyższymi i Grzybami; PZWL: Warszawa, Poland, 1984; p. 390. [Google Scholar]
- Żurek, G.; Wiewióra, B.; Żurek, M.; Łyszczarz, R. Environmental effect on Epichlöe endophyte occurrence and ergovaline concentration in wild populations of forage grasses in Poland. Plant Soil 2017, 410, 383–399. [Google Scholar] [CrossRef]
- Marczuk, J.; Ziętek, J.; Michalak, K.; Winiarczyk, S.; Lutnicki, K.; Brodzki, P.; Adaszek, Ł. Zatrucie ergowaliną w stadzie krów mlecznych—Opis przypadku. Med. Weter. 2018, 75, 635–639. [Google Scholar]
- Gibson, D.J. Grasses and Grassland Ecology; Oxford University Press: New York, NY, USA, 2009; pp. 1–305. [Google Scholar]
- Ileleji, K.E.; Sokhansanj, S.; Cundiff, J.S. Farm–Gate to Plant–Gate Deliver of Lignocellulosic Feedstocks from Plant Biomass for Biofuels Production; Blaschek, H., Ezeji, T.C., Scheffran, J., Eds.; Biofuels from Agricultural Wastes and Byproducts; Wiley-Blackwell: Hoboken, NJ, USA, 2010; pp. 117–159. [Google Scholar]
- Danielewicz, D.; Surma-Ślusarska, B.; Żurek, G.; Martyniak, D.; Kmiotek, M.; Dybka, K. Selected Grass Plants as Biomass Fuels and Raw Materials for Papermaking, Part II. Pulp and Paper Properties. Bioresources 2015, 4, 8552–8564. [Google Scholar] [CrossRef] [Green Version]
- Janiszewska, D.; Żurek, G.; Martyniak, D.; Prokopiuk, K. Comparison of the quality of particleboards made with addition of biomass from C3 and C4 grasses. Ind. Crop. Prod. 2021, submitted. [Google Scholar]
- Żurek, G. Use of amenity grasses—Status quo and innovations. Grassl. Sci. Eur. 2012, 17, 436–442. [Google Scholar]
- Prończuk, S.; Żurek, G.; Żyłka, D.; Prończuk, M.; Woś, M. Ocena śmiałka darniowego (Deschampsia cespitosa (L.) P.B.) w różnym użytkowaniu trawnikowym. Zesz. Probl. Post. Nauk Roln. Zesz. 2001, 474, 115–123. [Google Scholar]
- Żurek, G. Strzęplice (Koeleria sp.)—Mało znane gatunki do zadarniania terenów suchych. Cz. II. Ocena przydatności trawnikowej. Łąkarstwo W Polsce 2000, 3, 185–192. [Google Scholar]
- Żurek, G.; Sevcikova, M. Minor Grasses. In Handbook of Plant Breeding Vol. 5. Fodder Crops and Amenity Grasses; Boller, B., Veronesi, F., Posselt, U., Eds.; Springer + Businnes Media: Dordrecht, The Netherlands, 2010; pp. 381–394. [Google Scholar]
- Żurek, G.; Rybka, K.; Pogrzeba, M.; Krzyżak, J.; Prokopiuk, K. Chlorophyll a fluorescence in evaluation of the effect of heavy metal soil contamination on perennial grasses. PLoS ONE 2014, 9, e91. [Google Scholar] [CrossRef] [PubMed]
- Klimont, K.; Bulińska, Z. Sukcesja roślin na terenach poeksploatacyjnych kopalni siarki „Jeziórko”. Biul. Ihar 2010, 257/258, 29–37. [Google Scholar]
- Stefanek, P.; Sorbjan, P.; Stępień, M. Monitoring i jego wykorzystanie w eksploatacji i projektowaniu rozbudowy składowiska “Żelazny Most”. Infrastrukt. I Ekol. Teren. Wiej. 2010, 8, 105–116. [Google Scholar]
- Żurek, G.; Kochańska-Czembor, E.; Martyniak, D.; Prokopiuk, K. Doskonalenie nasiennictwa gatunków traw o niskiej rentowności na użytki i tereny zielone. Monogr. i Rozpr. Nauk. Ihar-Pib 2014, 48, 345–353. [Google Scholar]
- Falkowski, M. Trawy Polskie; PWRiL: Warszawa, Poland, 1982; p. 565. [Google Scholar]
- FAO. Voluntary Guidelines for the Conservation and Sustainable Use of Crop Wild Relatives and Wild Food Plants; Food and Agriculture Organization of the United Nations: Rome, Italy, 2017. [Google Scholar]
- Engels, J.M.; Thormann, I. Main Challenges and Actions Needed to Improve Conservation and Sustainable Use of Our Crop Wild Relatives. Plants 2020, 9, 968. [Google Scholar] [CrossRef]
- Podyma, W. Rola banku genów we wdrażaniu globalnych i krajowych planów ochrony różnorodności biologicznej. Biul. Ogrodów Bot. 2000, 9, 99–106. [Google Scholar]
- Krajowe Centrum Roślinnych Zasobów Genowych (NCPGR). Available online: http://bankgenow.edu.pl (accessed on 10 December 2020).
- Global Biodiversity Information Facility (GBIF). Available online: http://www.gbif.org (accessed on 10 November 2020).
- EURISCO Catalogue. Available online: http://eurisco.ipk–gatersleben.de (accessed on 10 November 2020).
- GENESYS Database. Available online: http://www.genesys–pgr.org (accessed on 10 December 2020).
- Meyer, A.; Barton, N. Botanic Gardens Are Important Contributors to Crop Wild Relative Preservation. Crop Sci. 2019, 59, 2404–2412. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Sharrock, S. Botanic gardens complement agricultural gene bank in collecting and conserving plant genetic diversity. Biopreserv. Biobanking 2018, 16, 384–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- KFGB—Kostrzyca Forest Gene Bank. Available online: https://www.lbg.lasy.gov.pl/ (accessed on 10 November 2020).






| Genus | No. of Species per Genus |
|---|---|
| Festuca | 33 |
| Poa | 19 |
| Bromus | 11 |
| Agrostis, Calamagrostis, Glyceria | 7 |
| Melica, Phleum, Stipa | 5 |
| Alopecurus, Koeleria, Trisetum | 4 |
| Agropyron, Deschampsia, Lolium | 3 |
| Anthoxanthum, Brachypodium, Dactylis, Digitaria, Elymus, Hierochloë, Holcus | 2 |
| Ammophila, Arrhenatherum, Avena, Briza, Corynephorus, Leersia, Milium, Molinia | |
| Phalaris, Phragmites | 1 |
| Variables | Correlation | |
|---|---|---|
| PC1 | PC2 | |
| Light value (L) | 0.785565 | 0.274023 |
| Soil moisture value (F) | −0.77331 | −0.34218 |
| Soil acidity (pH) value (R) | 0.601228 | −0.79816 |
| Taxon Name | Occurrence (Number of Preserved Specimens) from GBIF [92] | Number of Accessions from EURISCO [93] | Number of Accessions from NCPGR [91] |
|---|---|---|---|
| Festuca rubra | 46,617 | 3079 | 471 |
| F. arundinacea | 7947 | 2872 | 586 |
| F. gigantea | 3640 | 236 | 6 |
| F. tatrae | 55 | 4 | 1 |
| F. macutrensis | 6 | 0 | 0 |
| F. carpatica | 92 | 1 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dostatny, D.F.; Żurek, G.; Kapler, A.; Podyma, W. The Ex Situ Conservation and Potential Usage of Crop Wild Relatives in Poland on the Example of Grasses. Agronomy 2021, 11, 94. https://doi.org/10.3390/agronomy11010094
Dostatny DF, Żurek G, Kapler A, Podyma W. The Ex Situ Conservation and Potential Usage of Crop Wild Relatives in Poland on the Example of Grasses. Agronomy. 2021; 11(1):94. https://doi.org/10.3390/agronomy11010094
Chicago/Turabian StyleDostatny, Denise F., Grzegorz Żurek, Adam Kapler, and Wiesław Podyma. 2021. "The Ex Situ Conservation and Potential Usage of Crop Wild Relatives in Poland on the Example of Grasses" Agronomy 11, no. 1: 94. https://doi.org/10.3390/agronomy11010094
APA StyleDostatny, D. F., Żurek, G., Kapler, A., & Podyma, W. (2021). The Ex Situ Conservation and Potential Usage of Crop Wild Relatives in Poland on the Example of Grasses. Agronomy, 11(1), 94. https://doi.org/10.3390/agronomy11010094