

Distinguished Functions of Microglia in the Two Stages of Oxygen-Induced Retinopathy: A Novel Target in the Treatment of Ischemic Retinopathy

Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. PLX5622 Administration
2.3. Weight Monitor
2.4. Flow Cytometry
2.5. Staining
2.6. RT-PCR
2.7. ERG Examination
2.8. TUNEL Assay
2.9. Statistics
3. Results
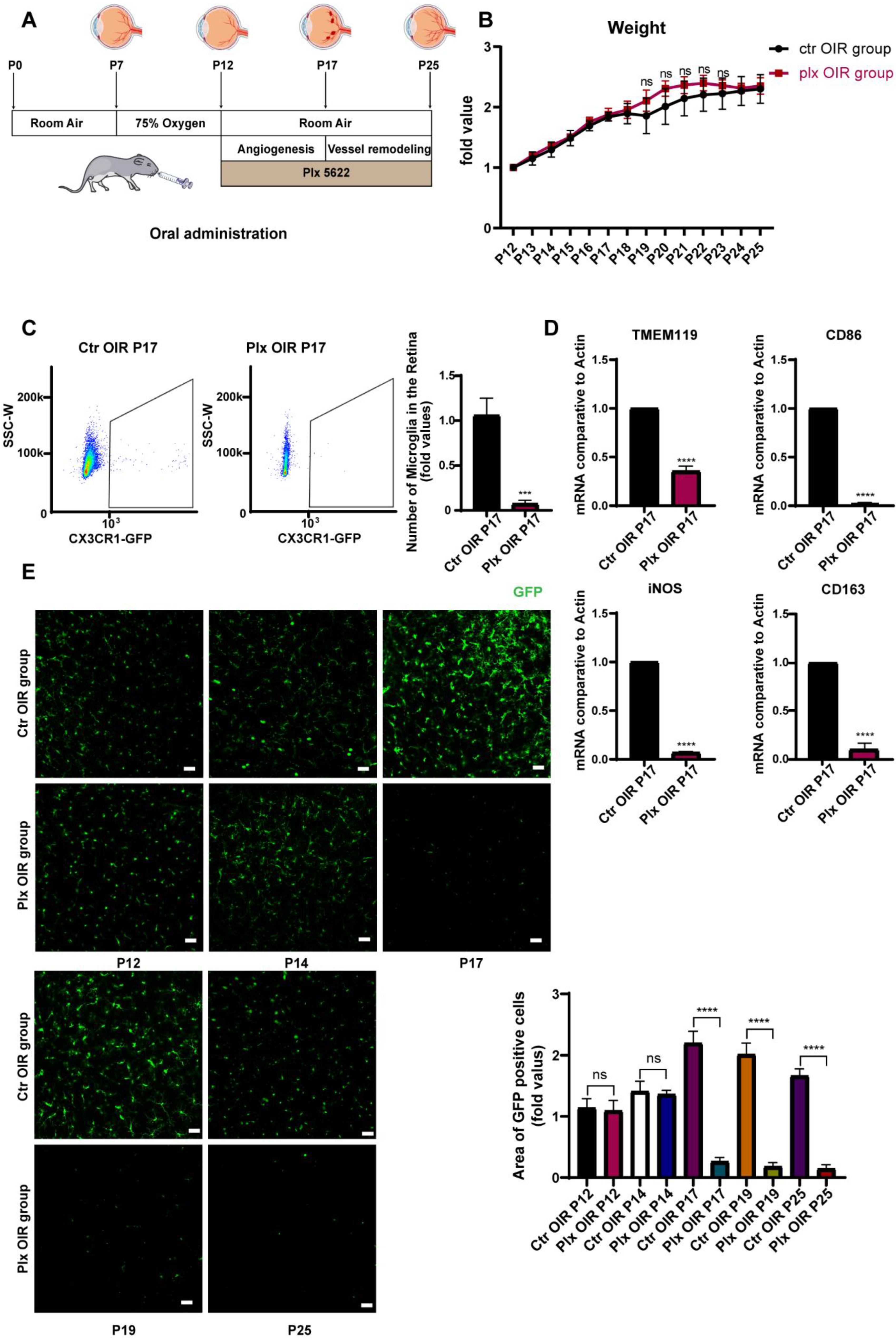
3.1. Plx5622 Effectively Removed Retinal Microglia without Affecting the Whole Conditions of Mice
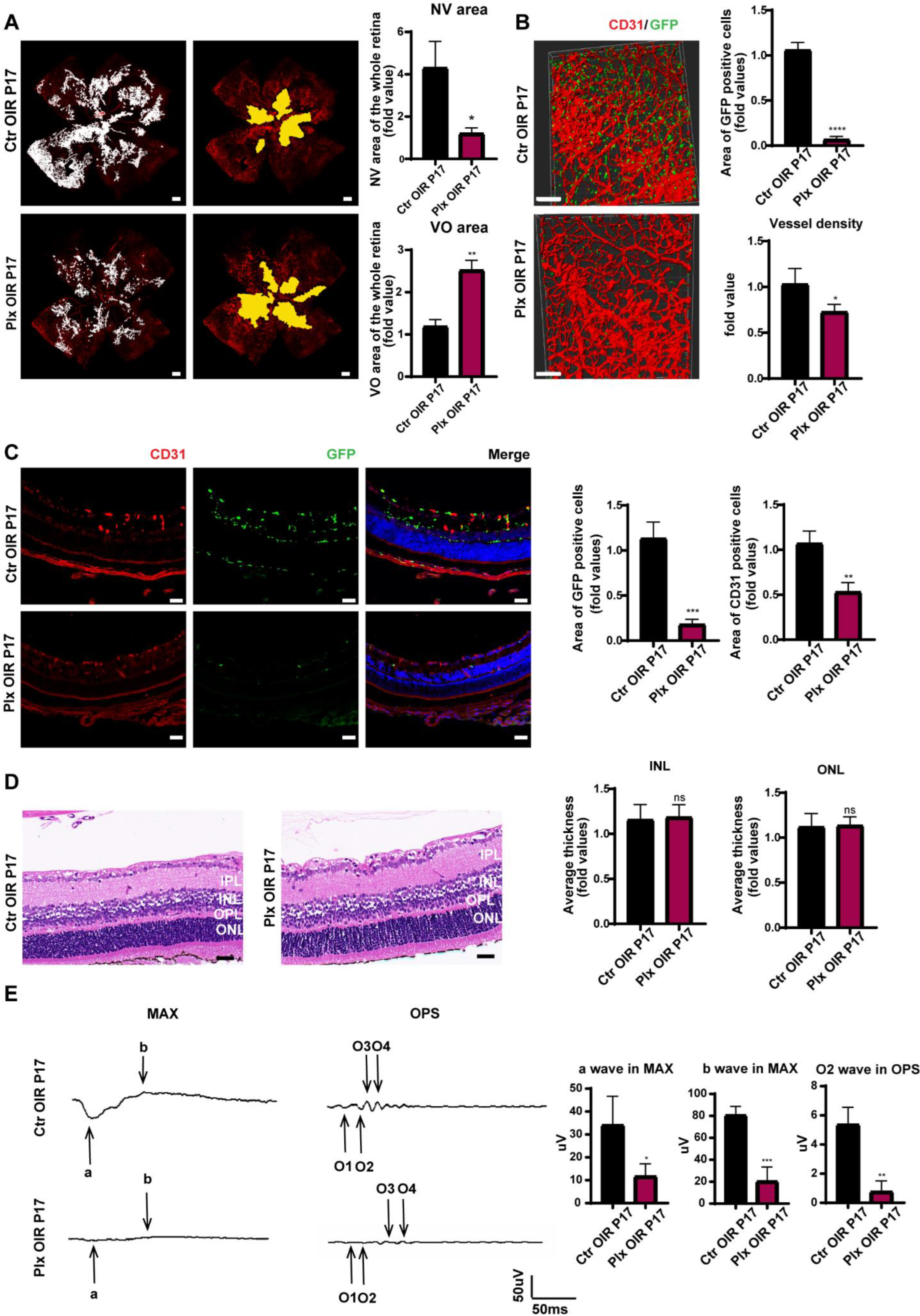
3.2. Elimination of Microglia Suppressed the Pathological Process in the Neovascularization Stage of OIR
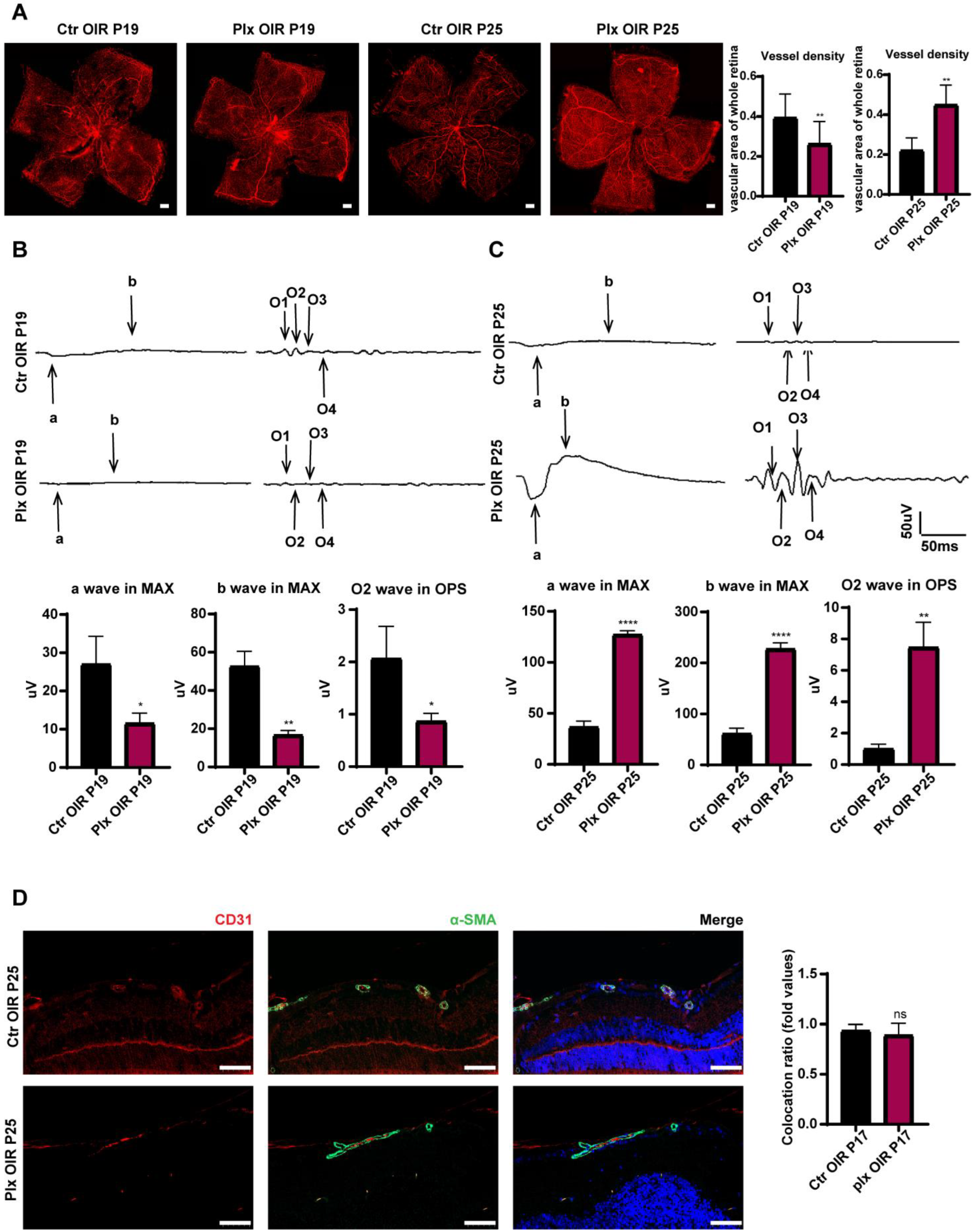
3.3. Elimination of Microglia Promoted the Repairing Process in Vascular Remodeling Stage of OIR
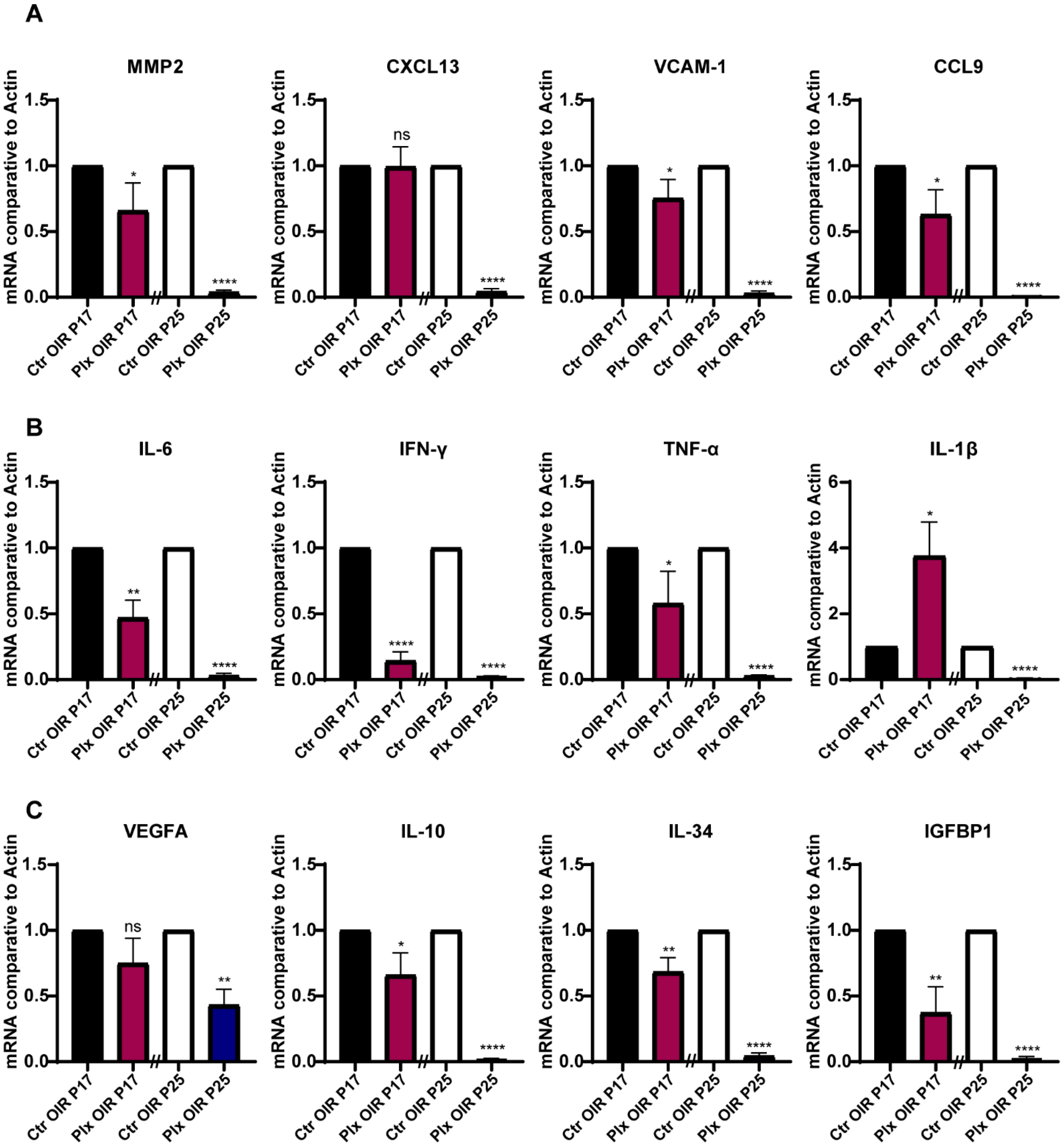
3.4. The Expression Profile of Cytokines in the Retina Declined Consistently after Microglia Removal
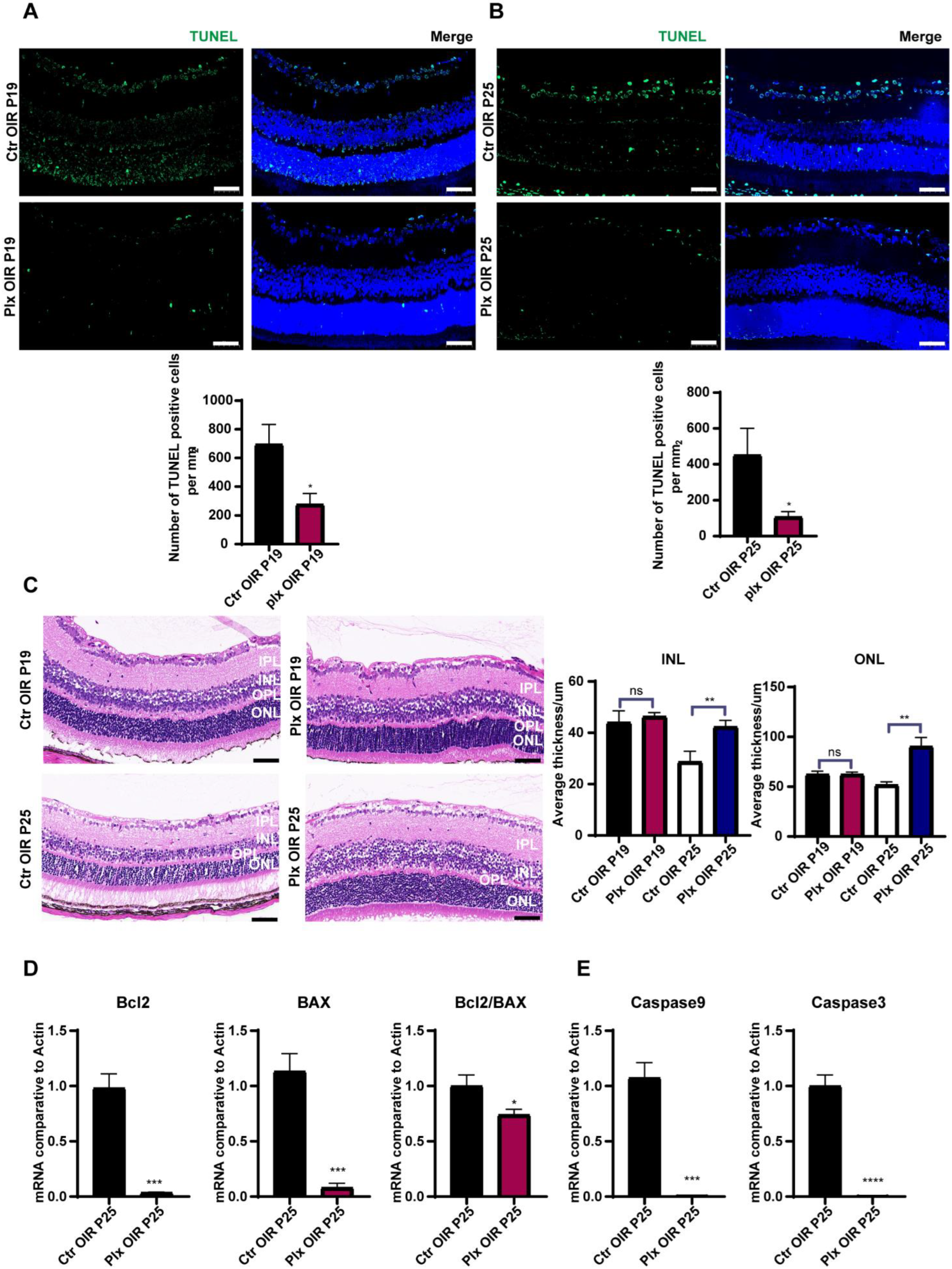
3.5. Microglia Played a Vital Role in Promoting Apoptosis in the Regression of OIR
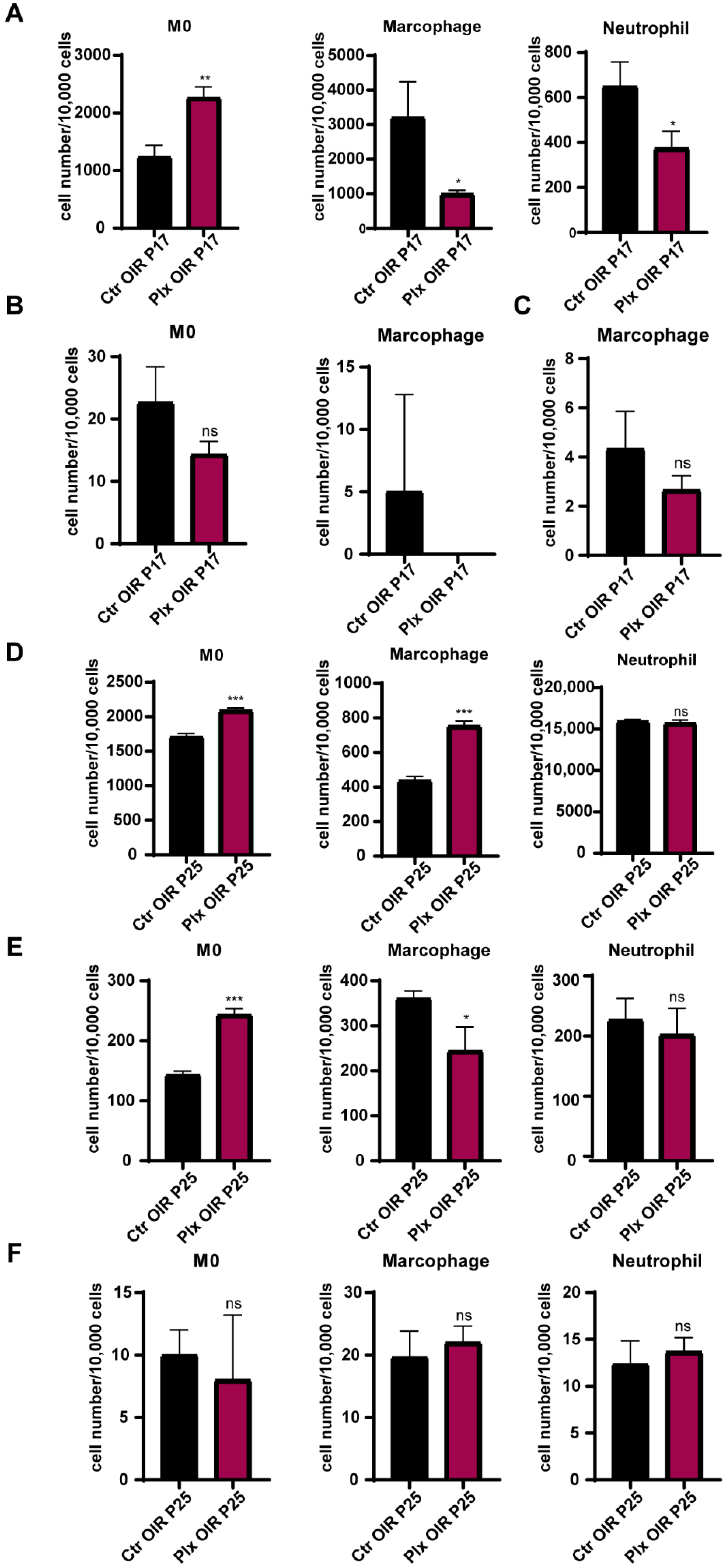
3.6. Depletion of Retinal Microglia Affected the Mobilization of Bone Marrow Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Binet, F.; Cagnone, G.; Crespo-Garcia, S.; Hata, M.; Neault, M.; Dejda, A.; Wilson, A.M.; Buscarlet, M.; Mawambo, G.T.; Howard, J.P.; et al. Neutrophil Extracellular Traps Target Senescent Vasculature for Tissue Remodeling in Retinopathy. Science 2020, 369, 6506. [Google Scholar] [CrossRef]
- Korn, C.; Augustin, H.G. Mechanisms of Vessel Pruning and Regression. Dev. Cell 2015, 34, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Mazzoni, J.; Smith, J.R.; Shahriar, S.; Cutforth, T.; Ceja, B.; Agalliu, D. The Wnt Inhibitor Apcdd1 Coordinates Vascular Remodeling and Barrier Maturation of Retinal Blood Vessels. Neuron 2017, 96, 1055–1069.e6. [Google Scholar] [CrossRef] [Green Version]
- Darlow, B.A.; Gilbert, C. Retinopathy of Prematurity—A World Update. Semin. Perinatol. 2019, 43, 315–316. [Google Scholar] [CrossRef]
- Antonetti, D.A.; Klein, R.; Gardner, T.W. Diabetic Retinopathy. N. Engl. J. Med. 2012, 366, 1227–1239. [Google Scholar] [CrossRef] [Green Version]
- Prinz, M.; Jung, S.; Priller, J. Microglia Biology: One Century of Evolving Concepts. Cell 2019, 179, 292–311. [Google Scholar] [CrossRef]
- Subhramanyam, C.S.; Wang, C.; Hu, Q.; Dheen, S.T. Microglia-Mediated Neuroinflammation in Neurodegenerative Diseases. Semin. Cell Dev. Biol. 2019, 94, 112–120. [Google Scholar] [CrossRef]
- Vecino, E.; Rodriguez, F.D.; Ruzafa, N.; Pereiro, X.; Sharma, S.C. Glia-Neuron Interactions in the Mammalian Retina. Prog. Retin. Eye Res. 2016, 51, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Arnold, T.; Betsholtz, C. The Importance of Microglia in the Development of the Vasculature in the Central Nervous System. Vasc. Cell 2013, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Checchin, D.; Sennlaub, F.; Levavasseur, E.; Leduc, M.; Chemtob, S. Potential Role of Microglia in Retinal Blood Vessel Formation. Invest. Ophthalmol. Vis. Sci. 2006, 47, 3595–3602. [Google Scholar] [CrossRef]
- Green, W.R. Clinicopathologic Studies of Treated Choroidal Neovascular Membranes. A Review and Report of Two Cases. Retina 1991, 11, 328–356. [Google Scholar] [CrossRef]
- Wang, S.K.; Cepko, C.L. Targeting Microglia to Treat Degenerative Eye Diseases. Front. Immunol. 2022, 13, 843558. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, W.; Chen, H.; Zhou, H.; Liu, Z.; Liu, Z.; Liu, Z.; Zhou, Y.; Zhou, X.; Yu, L.; et al. Sympathetic Nervous System Mediates Cardiac Remodeling after Myocardial Infarction in a Circadian Disruption Model. Front. Cardiovasc. Med. 2021, 8, 668387. [Google Scholar] [CrossRef]
- Ajami, B.; Bennett, J.L.; Krieger, C.; Tetzlaff, W.; Rossi, F. Local Self-Renewal Can Sustain Cns Microglia Maintenance and Function Throughout Adult Life. Nat. Neurosci. 2007, 10, 1538–1543. [Google Scholar] [CrossRef]
- Cserép, C.; Pósfai, B.; Dénes, Á. Shaping Neuronal Fate: Functional Heterogeneity of Direct Microglia-Neuron Interactions. Neuron 2021, 109, 222–240. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, Y.; Meng, J.; Wang, X.; Liu, X.; Li, W.; Zhou, Q.; Xiang, J.; Li, N.; Hou, S. Lgals3bp in Microglia Promotes Retinal Angiogenesis through Pi3k/Akt Pathway During Hypoxia. Invest. Ophthalmol. Vis. Sci. 2022, 63, 25. [Google Scholar] [CrossRef]
- Xu, Y.; Lu, X.; Hu, Y.; Yang, B.; Tsui, C.K.; Yu, S.; Lu, L.; Liang, X. Melatonin Attenuated Retinal Neovascularization and Neuroglial Dysfunction by Inhibition of Hif-1α-Vegf Pathway in Oxygen-Induced Retinopathy Mice. J. Pineal Res. 2018, 64, e12473. [Google Scholar] [CrossRef]
- Vessey, K.A.; Wilkinson-Berka, J.L.; Fletcher, E.L. Characterization of Retinal Function and Glial Cell Response in a Mouse Model of Oxygen-Induced Retinopathy. J. Comp. Neurol. 2011, 519, 506–527. [Google Scholar] [CrossRef]
- Connor, K.M.; Krah, N.M.; Dennison, R.J.; Aderman, C.M.; Chen, J.; Guerin, K.I.; Sapieha, P.; Stahl, A.; Willett, K.L.; Smith, L.E. Quantification of Oxygen-Induced Retinopathy in the Mouse: A Model of Vessel Loss, Vessel Regrowth and Pathological Angiogenesis. Nat. Protoc. 2009, 4, 1565–1573. [Google Scholar] [CrossRef] [Green Version]
- Dorrell, M.I.; Aguilar, E.; Jacobson, R.; Trauger, S.A.; Friedlander, J.; Siuzdak, G.; Friedlander, M. Maintaining Retinal Astrocytes Normalizes Revascularization and Prevents Vascular Pathology Associated with Oxygen-Induced Retinopathy. Glia 2010, 58, 43–54. [Google Scholar] [CrossRef]
- Spangenberg, E.; Severson, P.L.; Hohsfield, L.A.; Crapser, J.; Zhang, J.; Burton, E.A.; Zhang, Y.; Spevak, W.; Lin, J.; Phan, N.Y.; et al. Sustained Microglial Depletion with Csf1r Inhibitor Impairs Parenchymal Plaque Development in an Alzheimer’s Disease Model. Nat. Commun. 2019, 10, 3758. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Yin, J.; Sun, L.; Hu, Z.; Dou, G.; Zhang, Z.; Wang, H.; Guo, C.; Wang, Y. Impact of Minocycline on Vascularization and Visual Function in an Immature Mouse Model of Ischemic Retinopathy. Sci. Rep. 2017, 7, 7535. [Google Scholar] [CrossRef] [Green Version]
- Feldman, A.T.; Wolfe, D. Tissue Processing and Hematoxylin and Eosin Staining. Methods Mol. Biol. 2014, 1180, 31–43. [Google Scholar]
- Huang, Y.; Xu, Z.; Xiong, S.; Sun, F.; Qin, G.; Hu, G.; Wang, J.; Zhao, L.; Liang, Y.X.; Wu, T.; et al. Repopulated Microglia Are Solely Derived from the Proliferation of Residual Microglia after Acute Depletion. Nat. Neurosci. 2018, 21, 530–540. [Google Scholar] [CrossRef]
- Feng, X.; Valdearcos, M.; Uchida, Y.; Lutrin, D.; Maze, M.; Koliwad, S.K. Microglia Mediate Postoperative Hippocampal Inflammation and Cognitive Decline in Mice. JCI Insight 2017, 2, e91229. [Google Scholar] [CrossRef] [Green Version]
- Okunuki, Y.; Mukai, R.; Nakao, T.; Tabor, S.J.; Butovsky, O.; Dana, R.; Ksander, B.R.; Connor, K.M. Retinal Microglia Initiate Neuroinflammation in Ocular Autoimmunity. Proc. Natl. Acad. Sci. USA 2019, 116, 9989–9998. [Google Scholar] [CrossRef] [Green Version]
- Kokona, D.; Ebneter, A.; Escher, P.; Zinkernagel, M.S. Colony-Stimulating Factor 1 Receptor Inhibition Prevents Disruption of the Blood-Retina Barrier During Chronic Inflammation. J. Neuroinflammation 2018, 15, 340. [Google Scholar] [CrossRef] [Green Version]
- Lobov, I.B.; Cheung, E.; Wudali, R.; Cao, J.; Halasz, G.; Wei, Y.; Economides, A.; Lin, H.C.; Papadopoulos, N.; Yancopoulos, G.D.; et al. The Dll4/Notch Pathway Controls Postangiogenic Blood Vessel Remodeling and Regression by Modulating Vasoconstriction and Blood Flow. Blood 2011, 117, 6728–6737. [Google Scholar] [CrossRef]
- Ma, Y.; Li, T. Monitoring Dynamic Growth of Retinal Vessels in Oxygen-Induced Retinopathy Mouse Model. J. Vis. Exp. 2021, 170, e62410. [Google Scholar] [CrossRef]
- Huang, H. Pericyte-Endothelial Interactions in the Retinal Microvasculature. Int. J. Mol. Sci. 2020, 21, 7413. [Google Scholar] [CrossRef]
- Hong, S.; Beja-Glasser, V.F.; Nfonoyim, B.M.; Frouin, A.; Li, S.; Ramakrishnan, S.; Merry, K.M.; Shi, Q.; Rosenthal, A.; Barres, B.A.; et al. Complement and Microglia Mediate Early Synapse Loss in Alzheimer Mouse Models. Science 2016, 352, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Sellgren, C.M.; Gracias, J.; Watmuff, B.; Biag, J.D.; Thanos, J.M.; Whittredge, P.B.; Fu, T.; Worringer, K.; Brown, H.E.; Wang, J.; et al. Increased Synapse Elimination by Microglia in Schizophrenia Patient-Derived Models of Synaptic Pruning. Nat. Neurosci. 2019, 22, 374–385. [Google Scholar] [CrossRef] [PubMed]
- Thion, M.S.; Ginhoux, F.; Garel, S. Microglia and Early Brain Development: An Intimate Journey. Science 2018, 362, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Liu, M.Y.; Zhang, D.F.; Zhong, X.; Du, K.; Qian, P.; Yao, W.F.; Gao, H.; Wei, M.J. Baicalin Mitigates Cognitive Impairment and Protects Neurons from Microglia-Mediated Neuroinflammation Via Suppressing Nlrp3 Inflammasomes and Tlr4/Nf-Κb Signaling Pathway. CNS Neurosci. Ther. 2019, 25, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Boeck, M.; Thien, A.; Wolf, J.; Hagemeyer, N.; Laich, Y.; Yusuf, D.; Backofen, R.; Zhang, P.; Boneva, S.; Stahl, A.; et al. Temporospatial Distribution and Transcriptional Profile of Retinal Microglia in the Oxygen-Induced Retinopathy Mouse Model. Glia 2020, 68, 1859–1873. [Google Scholar] [CrossRef]
- O’Koren, E.G.; Yu, C.; Klingeborn, M.; Wong, A.Y.; Prigge, C.L.; Mathew, R.; Kalnitsky, J.; Msallam, R.A.; Silvin, A.; Kay, J.N.; et al. Microglial Function Is Distinct in Different Anatomical Locations During Retinal Homeostasis and Degeneration. Immunity 2019, 50, 723–737.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, S.R.; Roberts, J.M.; Zhang, J.; Steele, M.R.; Romero, C.O.; Bosco, A.; Vetter, M.L. Developmental Apoptosis Promotes a Disease-Related Gene Signature and Independence from Csf1r Signaling in Retinal Microglia. Cell Rep. 2019, 27, 2002–2013.e5. [Google Scholar] [CrossRef] [Green Version]
- Chitu, V.; Gokhan, Ş.; Nandi, S.; Mehler, M.F.; Stanley, E.R. Emerging Roles for Csf-1 Receptor and Its Ligands in the Nervous System. Trends Neurosci. 2016, 39, 378–393. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Xu, D.; Austin, C.D.; Caplazi, P.; Senger, K.; Sun, Y.; Jeet, S.; Young, J.; Delarosa, D.; Suto, E.; et al. Function of Csf1 and Il34 in Macrophage Homeostasis, Inflammation, and Cancer. Front. Immunol. 2019, 10, 2019. [Google Scholar] [CrossRef]
- Ebneter, A.; Kokona, D.; Jovanovic, J.; Zinkernagel, M.S. Dramatic Effect of Oral Csf-1r Kinase Inhibitor on Retinal Microglia Revealed by in Vivo Scanning Laser Ophthalmoscopy. Transl. Vis. Sci. Technol. 2017, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Funk, K.E.; Klein, R.S. Csf1r Antagonism Limits Local Restimulation of Antiviral Cd8(+) T Cells During Viral Encephalitis. J. Neuroinflammation 2019, 16, 22. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Given, K.S.; Dickson, E.L.; Owens, G.P.; Macklin, W.B.; Bennett, J.L. Concentration-Dependent Effects of Csf1r Inhibitors on Oligodendrocyte Progenitor Cells Ex Vivo and in Vivo. Exp. Neurol. 2019, 318, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Spiteri, A.G.; Ni, D.; Ling, Z.L.; Macia, L.; Campbell, I.L.; Hofer, M.J.; King, N.J. Plx5622 Reduces Disease Severity in Lethal Cns Infection by Off-Target Inhibition of Peripheral Inflammatory Monocyte Production. Front. Immunol. 2022, 13, 851556. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A.; Shinozaki, Y.; Kashiwagi, K.; Ohno, N.; Eto, K.; Wake, H.; Nabekura, J.; Koizumi, S. Microglia Mediate Non-Cell-Autonomous Cell Death of Retinal Ganglion Cells. Glia 2018, 66, 2366–2384. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic, J.; Liu, X.; Kokona, D.; Zinkernagel, M.S.; Ebneter, A. Inhibition of Inflammatory Cells Delays Retinal Degeneration in Experimental Retinal Vein Occlusion in Mice. Glia 2020, 68, 574–588. [Google Scholar] [CrossRef] [PubMed]
- Karlstetter, M.; Kopatz, J.; Aslanidis, A.; Shahraz, A.; Caramoy, A.; Linnartz-Gerlach, B.; Lin, Y.; Lückoff, A.; Fauser, S.; Düker, K.; et al. Polysialic Acid Blocks Mononuclear Phagocyte Reactivity, Inhibits Complement Activation, and Protects from Vascular Damage in the Retina. EMBO Mol. Med. 2017, 9, 154–166. [Google Scholar] [CrossRef]
- Shahraz, A.; Kopatz, J.; Mathy, R.; Kappler, J.; Winter, D.; Kapoor, S.; Schütza, V.; Scheper, T.; Gieselmann, V.; Neumann, H. Anti-Inflammatory Activity of Low Molecular Weight Polysialic Acid on Human Macrophages. Sci. Rep. 2015, 5, 16800. [Google Scholar] [CrossRef] [Green Version]
- Langmann, T.; Fauser, S. Polysialic Acid for Immunomodulation in an Animal Model for Wet Age-Related Macular Degeneration (Amd). Klin. Monbl. Augenheilkd. 2017, 234, 657–661. [Google Scholar]
- Schwarzer, P.; Kokona, D.; Ebneter, A.; Zinkernagel, M.S. Effect of Inhibition of Colony-Stimulating Factor 1 Receptor on Choroidal Neovascularization in Mice. Am. J. Pathol. 2020, 190, 412–425. [Google Scholar] [CrossRef] [Green Version]
- Foray, C.; Barca, C.; Winkeler, A.; Wagner, S.; Hermann, S.; Schaefers, M.; Grauer, O.M.; Zinnhardt, B.; Jacobs, A.H. In-terrogating Glioma-Associated Microglia/Macrophage Dynamics under Csf-1r Therapy with Multi-Tracer in Vivo Pet/Mr Imag-ing. J. Nucl. Med. 2022, 63, 1386–1393. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Catalog | Purpose |
|---|---|---|
| CD31 | BioLegend Cat# 102502 | IF |
| α-SMA | Abcam ab184705 | IF |
| CD45 | BioLegend Cat# 147716 | FACS |
| CD11b | BioLegend Cat# 101211 | FACS |
| Ly6C | BioLegend Cat# 128022 | FACS |
| Ly6G | BioLegend Cat# 127608 | FACS |
| F4/80 | BioLegend Cat# 123120 | FACS |
| Rhodopsin | Cell Signaling Cat# 21782 | IF |
| NeuN | Cell Signaling Cat# 24307 | IF |
| Glast | Proteintech Cat#20785-1-AP | IF |
| Caspase3 | Cell Signaling Cat# 9668 | IF |
| Name | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| Tmem119 | AGGAGTCCTGAGAGATTTGCGC | GTGGATTCGGACCAGTCTGA |
| CD86 | GTGGATTCGGAATTCAAGCTC | CAGCCAGTCTGAAGGCACA |
| iNOS | CATGATCGATCTTCGATATCGACG | TGACTGTAGCTAGCTATGCA |
| CD163 | GTACGTACCATCGATGCATGCAT | TGATCTGATGCATGCTAGTACG |
| MMP2 | CAAGGATGGACTCCTGGCACAT | CAAGGATGGACTCCTGGCACAT |
| CXCL13 | CATAGATCGGATTCAAGTTACGCC | GTAACCATTTGGCACGAGGATTC |
| VCAM-1 | GCTATGAGGATGGAAGACTCTGG | ACTTGTGCAGCCACCTGAGATC |
| CCL9 | TCCAGAGCAGTCTGAAGGCACA | CCGTGAGTTATAGGACAGGCAG |
| IL-6 | TACCACTTCACAAGTCGGAGGC | CTGCAAGTGCATCATCGTTGTTC |
| IFN-γ | CAGCAACAGCAAGGCGAAAAAGG | TTTCCGCTTCCTGAGGCTGGAT |
| TNF-α | GGTGCCTATGTCTCAGCCTCTT | GCCATAGAACTGATGAGAGGGAG |
| IL-1β | TGGACCTTCCAGGATGAGGACA | GTTCATCTCGGAGCCTGTAGTG |
| VEGFA | CTCTGGAACCTGAGACATCACC | AGGAGTCCTGAGAGATTTGCGC |
| Il-10 | CGGGAAGACAATAACTGCACCC | CGGTTAGCAGTATGTTGTCCAGC |
| Il-34 | ATCATCGGCATTTTGAACGAGGTC | ACCTTGGAAGCCCTACAGACGA |
| IGFBP1 | GCCCAACAGAAAGCAGGAGATG | GTAGACACACCAGCAGAGTCCA |
| Bcl2 | TGCATGTAACTACAGTACGACGTA | TGCATGACTCGTAGTCGATGCA |
| BAX | TGCATGCATAGCTAGTCGATGTCAT | TGCATATCGTAGCTGACAGCTG |
| Caspase3 | GGAGTCTGACTGGAAAGCCGAA | CTTCTGGCAAGCCATCTCCTCA |
| Caspase9 | GCTGTGTCAAGTTTGCCTACCC | CCAGAATGCCATCCAAGGTCTC |
| β-actin | GGCTGTATTCCCCTCCATCG | CCAGTTGGTAACAATGCCATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Z.; Jing, Y.; Niu, Y.; Chang, T.; Sun, J.; Guo, C.; Wang, Y.; Dou, G. Distinguished Functions of Microglia in the Two Stages of Oxygen-Induced Retinopathy: A Novel Target in the Treatment of Ischemic Retinopathy. Life 2022, 12, 1676. https://doi.org/10.3390/life12101676
Zhou Z, Jing Y, Niu Y, Chang T, Sun J, Guo C, Wang Y, Dou G. Distinguished Functions of Microglia in the Two Stages of Oxygen-Induced Retinopathy: A Novel Target in the Treatment of Ischemic Retinopathy. Life. 2022; 12(10):1676. https://doi.org/10.3390/life12101676
Chicago/Turabian StyleZhou, Ziyi, Yutong Jing, Yali Niu, Tianfang Chang, Jiaxing Sun, Changmei Guo, Yusheng Wang, and Guorui Dou. 2022. "Distinguished Functions of Microglia in the Two Stages of Oxygen-Induced Retinopathy: A Novel Target in the Treatment of Ischemic Retinopathy" Life 12, no. 10: 1676. https://doi.org/10.3390/life12101676
APA StyleZhou, Z., Jing, Y., Niu, Y., Chang, T., Sun, J., Guo, C., Wang, Y., & Dou, G. (2022). Distinguished Functions of Microglia in the Two Stages of Oxygen-Induced Retinopathy: A Novel Target in the Treatment of Ischemic Retinopathy. Life, 12(10), 1676. https://doi.org/10.3390/life12101676