Effects of Different Laying Hen Species on Odour Emissions

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
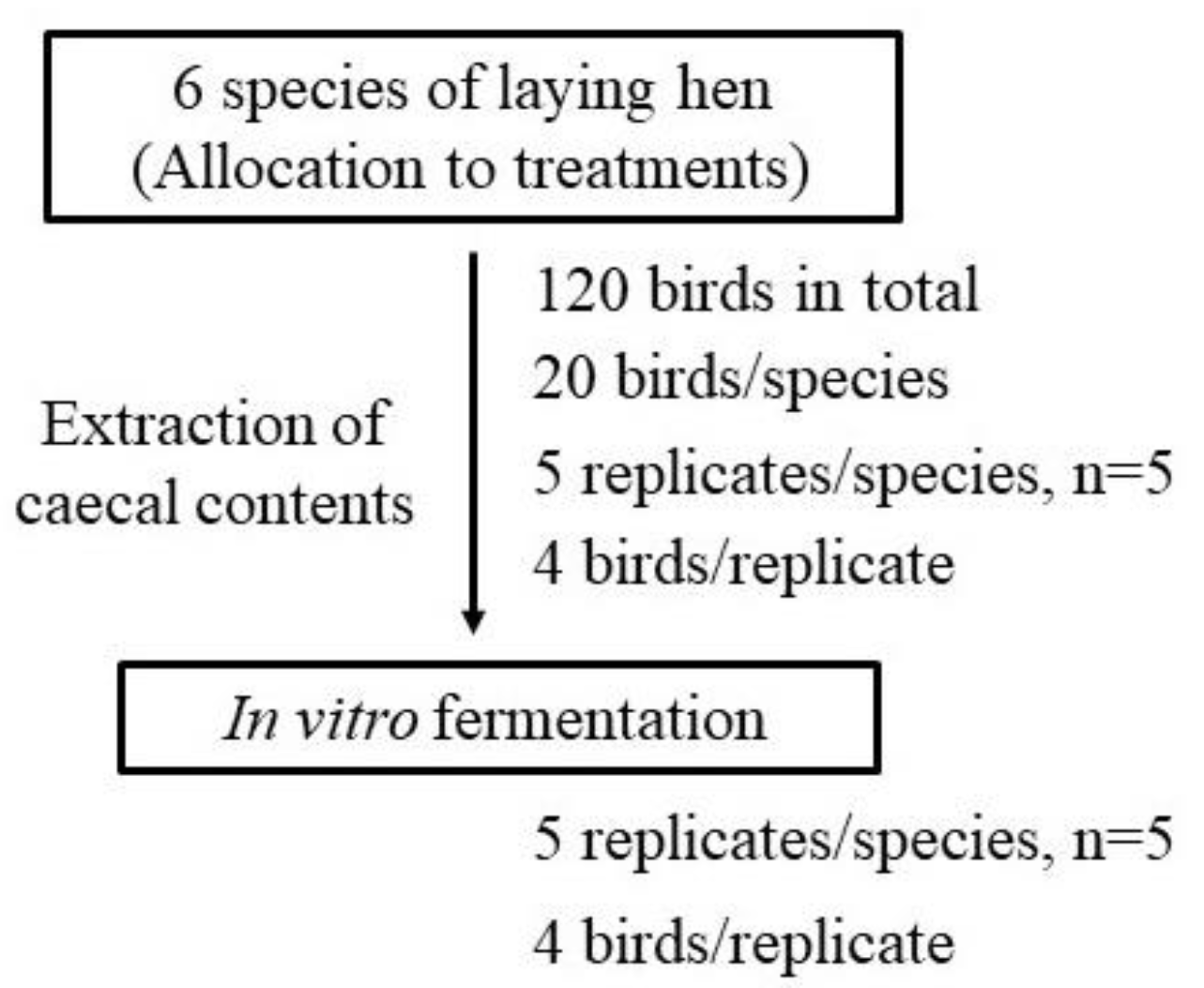
2.2. Animal Management and Preparation of Inoculums
2.3. In Vitro Fermentation and Sample Collection
2.4. Odour Gas Measurements
2.5. Analysis of Fermentation Liquid Samples
2.6. DNA Extraction and aprA and ureC Gene Quantification
2.7. Statistical Analysis
3. Results
3.1. Odour Gas Production
3.2. Production Performance
3.3. VFA Production in the Laying Hen Caecum
3.4. Biochemical Indexes in Fermentation Liquid among Different Species
3.5. Changes in Sulphate (SO42−) Concentration in the Caecum
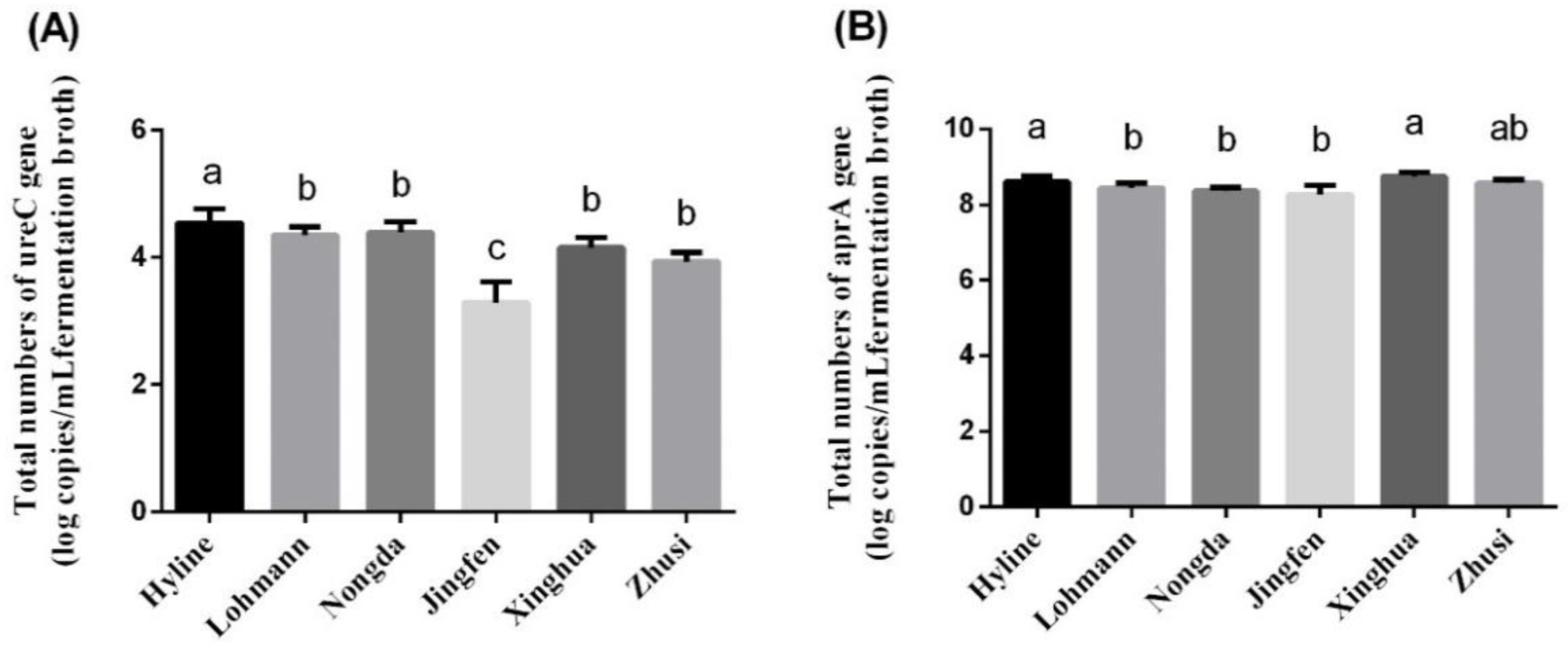
3.6. Quantitation of the Gene UreC and Gene AprA
3.7. Correlation of Odour Emission with Intestinal Activities, Microbial Genes and Production Performance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wu, Y.; Qin, F. Analysis on production efficiency of laying hens in China—Based on the survey data of five provinces. J. Agric. Sci. Camb. 2019, 9, 280–286. [Google Scholar]
- Wi, J.; Lee, S.; Kim, E.; Lee, M.; Koziel, J.; Ahn, H. Evaluation of semi-continuous pit manure recharge system performance on mitigation of ammonia and hydrogen sulfide emissions from a swine finishing barn. Atmosphere 2019, 10, 170. [Google Scholar] [CrossRef] [Green Version]
- Loftus, C.; Afsharinejad, Z.; Sampson, P.; Vedal, S.; Torres, E.; Arias, G.; Tchong-French, M.; Karr, C. Estimated time-varying exposures to air emissions from animal feeding operations and childhood asthma. Int. J. Hyg. Environ. Health 2020, 223, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Heinzen, T. Recent developments in the quantification and regulation of air emissions from animal feeding operations. Curr. Environ. Health Rep. 2015, 2, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.F.; Liu, Y.; Murphy, J.; Maghirang, R. Ammonia and methane emission factors from cattle operations expressed as losses of dietary nutrients or energy. Agriculture 2017, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, A.C.; Predicala, B.Z. Occupational exposure risk for swine workers in confined housing facilities. J. Agric. Saf. Health 2019, 25, 37–50. [Google Scholar] [CrossRef]
- Lin, X.J.; Cortus, E.L.; Zhang, R.; Jiang, S.; Heber, A.J. Ammonia, hydrogen sulfide, carbon dioxide and particulate matter emissions from California high-rise layer houses. Atmos. Environ. 2012, 46, 81–91. [Google Scholar] [CrossRef]
- Naseem, S.; King, A.J. Ammonia production in poultry houses can affect health of humans, birds, and the environment—Techniques for its reduction during poultry production. Environ. Sci. Pollut. Res. 2018, 25, 15269–15293. [Google Scholar] [CrossRef]
- Shi, Q.; Wang, W.; Chen, M.; Zhang, H.; Xu, S. Ammonia induces Treg/Th1 imbalance with triggered NF-κB pathway leading to chicken respiratory inflammation response. Sci. Total. Environ. 2019, 659, 354–362. [Google Scholar] [CrossRef]
- Wang, Y.; Xue, W.T.; Zhu, Z.P.; Yang, J.F.; Li, X.R.; Tian, Z.; Dong, H.M.; Zou, G.Y. Mitigating ammonia emissions from typical broiler and layer manure management–A system analysis. Waste Manag. 2019, 93, 23–33. [Google Scholar] [CrossRef]
- Malone Rubright, S.L.; Pearce, L.L.; Peterson, J. Environmental toxicology of hydrogen sulfide. Nitric Oxide 2017, 71, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.Y.; Chi, Q.R.; Wang, D.X.; Chi, X.; Teng, X.H.; Li, S. Hydrogen sulfide inhalation-induced immune damage is involved in oxidative stress, inflammation, apoptosis and the Th1/Th2 imbalance in broiler bursa of Fabricius. Ecotoxicol. Environ. Saf. 2018, 164, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.Z.; Zhang, T.; Lin, H.J.; Chang, Y.; Xing, J. Role of hydrogen sulfide on autophagy in liver injuries induced by selenium deficiency in chickens. Bio. Trace Elem. Res. 2017, 175, 194–203. [Google Scholar]
- Chi, Q.R.; Wang, D.X.; Hu, X.Y.; Li, S.P.; Li, S. Hydrogen sulfide gas exposure induces necroptosis and promotes inflammation through the MAPK/NF-κB pathway in broiler spleen. Oxidative. Med. Cell. Longev. 2019, 2019, 8061823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, C.H.; Ogden, J.M.; Pohl, H.R.; Scinicariello, F.; Ingerman, L.; Barber, L.; Citra, M.J. Toxicological Profile for Hydrogen Sulfide and Carbonyl Sulfide. Agency for Toxic Substances and Disease Registry. Division of Toxicology and Human Health Sciences; Environmental Toxicology Branch: Atlanta, GA, USA, 2016. [Google Scholar]
- Hofmann, T.; Schmucker, S.S.; Bessei, W.; Grashorn, M.; Stefanski, V. Impact of housing environment on the immune system in chickens: A review. Animals 2020, 10, 1138. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.S.; Ijssennagger, N.; Kies, A.K.; van Mil, S.W.C. Protein fermentation in the gut implications for intestinal dysfunction in humans, pigs, and poultry. Am. J. Physiol.-Gastrointest. Live Physiol. 2018, 315, G159–G170. [Google Scholar] [CrossRef]
- Wang, A.P.; Wang, Y.; Liao, X.D.; Wu, Y.B.; Liang, J.B.; Laudadio, V.; Tufarelli, V. Sodium butyrate mitigates in vitro ammonia generation in cecal content of laying hens. Environ. Sci. Pollut. R. 2016, 23, 16272–16279. [Google Scholar] [CrossRef]
- Zhang, M.L.; Wang, H.X. Organic wastes as carbon sources to promote sulfate reducing bacterial activity for biological remediation of acid mine drainage. Miner. Eng. 2014, 69, 81–90. [Google Scholar] [CrossRef]
- Kushkevych, I.; Leščanová, O.; Dordević, D.; Jančíková, S.; Hošek, J.; Vítězová, M.; Buňková, L.; Drago, L. The sulfate-reducing microbial communities and meta-analysis of their occurrence during diseases of small–large intestine axis. J. Clin. Med. 2019, 8, 1656. [Google Scholar] [CrossRef] [Green Version]
- Washio, J.; Shimada, Y.; Yamada, M.; Sakamaki, R.; Takahashi, N. Effects of pH and lactate on hydrogen sulfide production by oral Veillonella spp. Appl. Environ. Microb. 2014, 80, 4184–4188. [Google Scholar] [CrossRef] [Green Version]
- Snopková, K.; Sedlář, K.; Bosák, J.; Chaloupková, E.; Sedláček, I.; Provazník, I.; Šmajs, D. Free-Living Enterobacterium Pragia fontium 24613: Complete genome sequence and metabolic profiling. Evol. Bioinform. 2017, 13, 1176934317700863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Van Treuren, W.; Knight, R.; Bell, J.T.; et al. Human Genetics Shape the Gut Microbiome. Cell 2014, 159, 789–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, D.; Tiezzi, F.; Schillebeeckx, C.; McNulty, N.P.; Schwab, C.; Shull, C.; Maltecca, C. Host contributes to longitudinal diversity of fecal microbiota in swine selected for lean growth. Microbiome 2018, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, S. Effects of laying-hen strain on manure properties and ammonia emission. Trans. ASABE 2012, 55, 1059–1065. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Chen, W.L.; Tang, S.G.H.; Jahromi, M.F.; Candyrine, S.C.L.; Idrus, Z.; Abdullah, N.; Liang, J.B. Metagenomics analysis reveals significant modulation of cecal microbiota of broilers fed palm kernel expeller diets. Poult. Sci. 2019, 98, 56–68. [Google Scholar] [CrossRef]
- Menke, K.H.; Steingass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Deng, Y.F.; Liao, X.D.; Wang, Y.; Liang, J.B.; Tufarelli, V. Prebiotics mitigate in vitro sulfur-containing odour generation in caecal content of pigs. Ital. J. Anim. Sci. 2015, 14, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Guan, S.Y. Soil Enzyme and Its Research Methods; Agriculture Press: Beijing, China, 1986. [Google Scholar]
- Wall, E.; Simm, G.; Moran, D. Developing speciesing schemes to assist mitigation of greenhouse gas emissions. Animal 2010, 4, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.L.; Long, W.T.; Chadwick, D.; Velthof, G.L.; Oenema, O.; Ma, W.Q.; Wang, J.J.; Qin, W.; Hou, Y.; Zhang, F.S. Can dietary manipulations improve the productivity of pigs with lower environmental and economic cost? A global meta-analysis. Agric. Ecosyst. Environ. 2020, 289, 106748. [Google Scholar] [CrossRef]
- Herrero, M.; Havlík, P.; Valin, H.; Notenbaert, A.; Rufino, M.C.; Thornton, P.K.; Blümmel, M.; Weiss, F.; Grace, D.; Obersteiner, M. Biomass use, production, feed efficiencies, and greenhouse gas emissions from global livestock systems. Proc. Natl. Acad. Sci. USA 2013, 110, 20888–20893. [Google Scholar] [CrossRef] [Green Version]
- Ngwabie, N.M.; Acobta, A.N.; Manga, V.E.; VanderZaag, A.C. Emission factors of greenhouse gases from layer and broiler barns in Cameroon. Atmos. Environ. 2018, 176, 54–59. [Google Scholar] [CrossRef]
- Steenfeldt, S.; Hammershøj, M. Organic egg production. I: Effects of different dietary protein contents and forage material on organic egg production, nitrogen and mineral retention and total tract digestibility of nutrients of two hen genotypes. Animal Feed Sci. Technol. 2015, 209, 186–201. [Google Scholar] [CrossRef]
- Wu, C.D.; Yang, F.; Brancher, M.; Liu, J.M.; Qu, C.; Piringer, M.; Schauberger, G. Determination of ammonia and hydrogen sulfide emissions from a commercial dairy farm with an exercise yard and the health-related impact for residents. Environ. Sci. Pollut. Res. 2020, 27, 37684–37698. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Kim, I.H. The effects of betaine supplementation in diets containing different levels of crude protein and methionine on the growth performance, blood components, total tract nutrient digestibility, excreta noxious gas emission, and meat quality of the broiler chickens. Poult. Sci. 2019, 98, 6808–6815. [Google Scholar] [PubMed]
- Magee, E.A.; Richardson, C.J.; Hughes, R.; Cummings, J.H. Contribution of dietary protein to sulfide production in the large intestine: An in vitro and a controlled feeding study in humans. Am. J. Clin. Nutr. 2000, 72, 1488–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Recharla, N.; Kim, K.; Park, J.; Jeong, J.; Jeong, Y.; Lee, H.; Hwang, O.; Ryu, J.; Baek, Y.; Oh, Y.; et al. Effects of amino acid composition in pig diet on odorous compounds and microbial characteristics of swine excreta. J. Anim. Sci. Technol. 2017, 59, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, J.D.; Chen, X.; Liao, X.D. Screening of single or combined administration of 9 probiotics to reduce ammonia emissions from laying hens. Poult. Sci. 2019, 98, 3977–3988. [Google Scholar] [CrossRef]
- Nguyen, Q.H.; Le, P.D.; Chim, C.; Le, N.D.; Fievez, V. Potential to mitigate ammonia emission from slurry by increasing dietary fermentable fiber through inclusion of tropical byproducts in practical diets for growing pigs. Asian Austral. J. Anim. 2019, 32, 574–584. [Google Scholar] [CrossRef]
- Le, P.D.; Aarnink, A.J.; Ogink, N.W. Odour from animal production facilities: Its relationship to diet. Nutr. Res. Rev. 2005, 18, 3–30. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, C.J.; Lynch, M.B.; Callan, J.J.; O’Doherty, J.V. Dietary supplementation with chitosan at high and low crude protein concentrations promotes Enterobacteriaceae in the cecum and colon and increases manure odour emissions from finisher boars. Livest. Sci. 2010, 134, 198–201. [Google Scholar] [CrossRef]
- Xu, Y.; Tian, Y.; Cao, Y.; Li, J.; Guo, H.; Su, Y.; Tian, Y.; Wang, C.; Wang, T.; Zhang, L. Probiotic properties of Lactobacillus paracasei subsp. paracasei L1 and its growth performance-promotion in chicken by improving the intestinal microflora. Front. Physiol. 2019, 10, 937. [Google Scholar] [CrossRef] [PubMed]
- Al-Khalaifa, H.; Al-Nasser, A.; Al-Surayee, T.; Al-Kandari, S.; Al-Enzi, N.; Al-Sharrah, T.; Ragheb, G.; Al-Qalaf, S.; Mohammed, A. Effect of dietary probiotics and prebiotics on the performance of broiler chickens. Poult. Sci. 2019, 98, 4465–4479. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.L.; Hornig, M.; Parekh, T.; Lipkin, W.I. Application of novel PCR-based methods for detection, quantitation, and phylogenetic characterization of Sutterella species in intestinal biopsy samples from children with autism and gastrointestinal disturbances. Mbio 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandit, R.J.; Hinsu, A.T.; Patel, N.V.; Koringa, P.G.; Jakhesara, S.J.; Thakkar, J.R.; Shah, T.M.; Limon, G.; Psifidi, A.; Guitian, J.; et al. Microbial diversity and community composition of caecal microbiota in commercial and indigenous Indian chickens determined using 16s rDNA amplicon sequencing. Microbiome 2018, 6, 115. [Google Scholar] [CrossRef]
- Jin, D.; Zhao, S.G.; Wang, P.P.; Zheng, N.; Bu, D.P.; Beckers, Y.; Wang, J.Q. Insights into abundant rumen ureolytic bacterial community using rumen simulation system. Front. Microbiol. 2016, 7, 1006. [Google Scholar] [CrossRef] [Green Version]
- Keikha, M.; Eslami, M.; Yousefi, B.; Ghasemian, A.; Karbalaei, M. Potential antigen candidates for subunit vaccine development against Helicobacter pylori infection. J. Cell. Physiol. 2019, 234, 21460–21470. [Google Scholar] [CrossRef]
- Miller, W.G.; Yee, E.; Chapman, M.H.; Bono, J.L. Comparative genomics of all three campylobacter sputorum biovars and a novel cattle-associated C. sputorum clade. Genome Biol. Evol. 2017, 9, 1513–1518. [Google Scholar] [CrossRef] [Green Version]
- Eschenlauer, S.C.P.; McKain, N.; Walker, N.D.; McEwan, N.R.; Newbold, C.J.; Wallace, R.J. Ammonia production by ruminal microorganisms and enumeration, isolation, and characterization of bacteria capable of growth on peptides and amino acids from the sheep rumen. Appl. Environ. Microb. 2002, 68, 4925–4931. [Google Scholar] [CrossRef] [Green Version]
- Stern, R.A.; Mozdziak, P.E. Glutamine synthetase in avian muscle contributes to a positive myogenic response to ammonia compared with mammalian muscle. Am. J. Physiol.-Regul. I. 2019, 317, R214–R221. [Google Scholar] [CrossRef]
- Stern, R.A.; Mozdziak, P.E. Differential ammonia metabolism and toxicity between avian and mammalian species, and effect of ammonia on skeletal muscle: A comparative review. J. Anim. Physiol. Anim. Nutr. 2019, 103, 774–785. [Google Scholar] [CrossRef] [Green Version]
- Kushkevych, I.; Kos, J.; Kollar, P.; Kralova, K.; Jampilek, J. Activity of ring-substituted 8-hydroxyquinoline-2-carboxanilides against intestinal sulfate-reducing bacteria Desulfovibrio piger. Med. Chem. Res. 2018, 27, 278–284. [Google Scholar] [CrossRef]
- Meyer, B.; Kuever, J. Molecular analysis of the distribution and phylogeny of dissimilatory adenosine-5’-phosphosulfate reductase-encoding genes (aprBA) among sulfur-oxidizing prokaryotes. Microbiology 2007, 15, 3478–3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telman, W.; Dietz, K.J. Thiol redox-regulation for efficient adjustment of sulfur metabolism in acclimation to abiotic stress. J. Exp. Bot. 2019, 70, 4223–4236. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Total Volume (mL) | Concentration of NH3 (µg/L) | Concentration of H2S (µg/L) | Total NH3 Emission (µg) | Total H2S Emission (µg) | NH3 + H2S Emission (µg) |
|---|---|---|---|---|---|---|
| Hyline | 31.20 c | 2404.87 a | 551.43 a | 75.15 a | 17.30 a | 92.45 a |
| Lohmann | 25.88 d | 1720.54 c | 347.41 bc | 44.47 c | 8.92 b | 53.39 c |
| Nongda | 33.90 bc | 2032.32 b | 521.93 ab | 68.96 ab | 17.44 a | 86.40 b |
| Jingfen | 39.11 a | 1735.99 c | 492.63 bc | 67.86 b | 17.65 a | 85.42 b |
| Xinghua | 36.62 ab | 772.57 d | 318.41 c | 28.24 d | 11.70 b | 39.94 c |
| Zhusi | 36.49 ab | 1952.34 b | 528.88 ab | 71.24 ab | 19.09 a | 90.33 a |
| SEM | 0.02 | 30.60 | 56.81 | 2.18 | 1.71 | 2.57 |
| p-value | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Species | ADFI (g/day) | Egg Production (%) | Egg Weight (g) | FCR (g of Feed: g of Egg) |
|---|---|---|---|---|
| Hyline | 112.87 b | 97.22 a | 53.53 a | 2.17 e |
| Lohmann | 120.13 a | 96.34 a | 53.52 a | 2.33 d |
| Nongda | 91.44 e | 83.96 c | 44.04 c | 2.47 c |
| Jingfen | 104.99 c | 91.89 b | 50.51 b | 2.26 d |
| Xinghua | 90.58 e | 75.34 d | 41.04 e | 2.93 b |
| Zhusi | 96.11 d | 70.06 e | 42.59 d | 3.22 a |
| SEM | 0.53 | 0.39 | 0.29 | 0.02 |
| p-value | <0.01 | <0.01 | <0.01 | <0.01 |
| Species | Acetic Acid (mmol/mL) | Propionic Acid (mmol/mL) | Isobutyric Acid (mmol/mL) | Butyrate Acid (mmol/mL) | Isovaleric Acid (mmol/mL) | Valeric Acid (mmol/mL) | Total VFAs (mmol/mL) |
|---|---|---|---|---|---|---|---|
| Hyline | 49.12 a | 7.30 b | 0.71 b | 9.05 a | 1.96 b | 1.26 b | 69.47 a |
| Lohmann | 38.29 b | 5.87 d | 0.57 b | 7.52 b | 1.35 d | 0.91 c | 54.51 b |
| Nongda | 23.40 c | 3.61 e | 0.56 b | 5.49 c | 0.88 e | 0.56 d | 34.49 c |
| Jingfen | 41.48 b | 6.50 c | 0.65 b | 7.93 b | 1.70 c | 1.15 b | 59.41 b |
| Xinghua | 17.44 d | 2.69 f | 0.24 c | 4.98 c | 0.49 f | 0.35 e | 26.20 d |
| Zhusi | 52.13 a | 8.36 a | 1.39 a | 9.14 a | 2.46 a | 1.52 a | 75.00 a |
| SEM | 1.08 | 0.15 | 0.06 | 0.16 | 0.06 | 0.03 | 1.45 |
| p-value | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Species | pH | Urease Activity (mg/mL) | Ammonium Nitrogen (mg/mL) | Uric Acid (mmol/L) | Urea (mmol/L) |
|---|---|---|---|---|---|
| Hyline | 7.72 bc | 0.20 a | 0.46 a | 0.31 a | 0.31 ab |
| Lohmann | 7.76 b | 0.27 a | 0.44 a | 0.35 a | 0.40 ab |
| Nongda | 7.65 c | 0.18 b | 0.46 a | 0.36 a | 0.44 a |
| Jingfen | 7.54 d | 0.12 b | 0.39 a | 0.17 b | 0.26 bc |
| Xinghua | 8.06 a | 0.15 b | 0.28 b | 0.12 b | 0.16 c |
| Zhusi | 7.64 cd | 0.13 b | 0.46 a | 0.16 b | 0.34 ab |
| SEM | 0.03 | 0.02 | 0.03 | 0.02 | 0.04 |
| p-value | <0.01 | 0.01 | 0.01 | <0.01 | <0.01 |
| Gas Production | pH | Urease Activity (mg/mL) | Ammonium Nitrogen (mg/mL) | Uric Acid (mmol/L) | Number of ureC Genes | Reduction of SO42− | Number of aprA Genes |
|---|---|---|---|---|---|---|---|
| Total volume (mL) | −0.43 | −0.94 ** | −0.37 | −0.94 ** | −0.77 † | 0.001 | 0.20 |
| Concentration of NH3 (µg/L) | −0.33 | 0.03 | 0.93 ** | 0.67 | 0.47 | N/A | N/A |
| Total NH3 emission (µg) | −0.89 * | −0.77 † | 0.90 ** | 0.47 | −0.33 | N/A | N/A |
| Concentration of H2S (µg/L) | −0.48 | N/A | N/A | N/A | N/A | 0.81 * | −0.46 |
| Total H2S emission (µg) | −0.48 | N/A | N/A | N/A | N/A | 0.49 | −0.35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, D.; Mi, J.; Wu, Y.; Liang, J.; Liao, X.; Wang, Y. Effects of Different Laying Hen Species on Odour Emissions. Animals 2020, 10, 2172. https://doi.org/10.3390/ani10112172
Lu D, Mi J, Wu Y, Liang J, Liao X, Wang Y. Effects of Different Laying Hen Species on Odour Emissions. Animals. 2020; 10(11):2172. https://doi.org/10.3390/ani10112172
Chicago/Turabian StyleLu, Dongdong, Jiandui Mi, Yinbao Wu, Juanboo Liang, Xindi Liao, and Yan Wang. 2020. "Effects of Different Laying Hen Species on Odour Emissions" Animals 10, no. 11: 2172. https://doi.org/10.3390/ani10112172
APA StyleLu, D., Mi, J., Wu, Y., Liang, J., Liao, X., & Wang, Y. (2020). Effects of Different Laying Hen Species on Odour Emissions. Animals, 10(11), 2172. https://doi.org/10.3390/ani10112172