Ruminal Methanogenic Responses to the Thiamine Supplementation in High-Concentrate Diets

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Substrates and Experimental Design
2.2. Fermentation Parameters Measurement
2.3. Metagenomic Sequencing Process
2.4. Statistical Analysis
3. Results
3.1. Rumen Fermentation Parameters and Methane Production
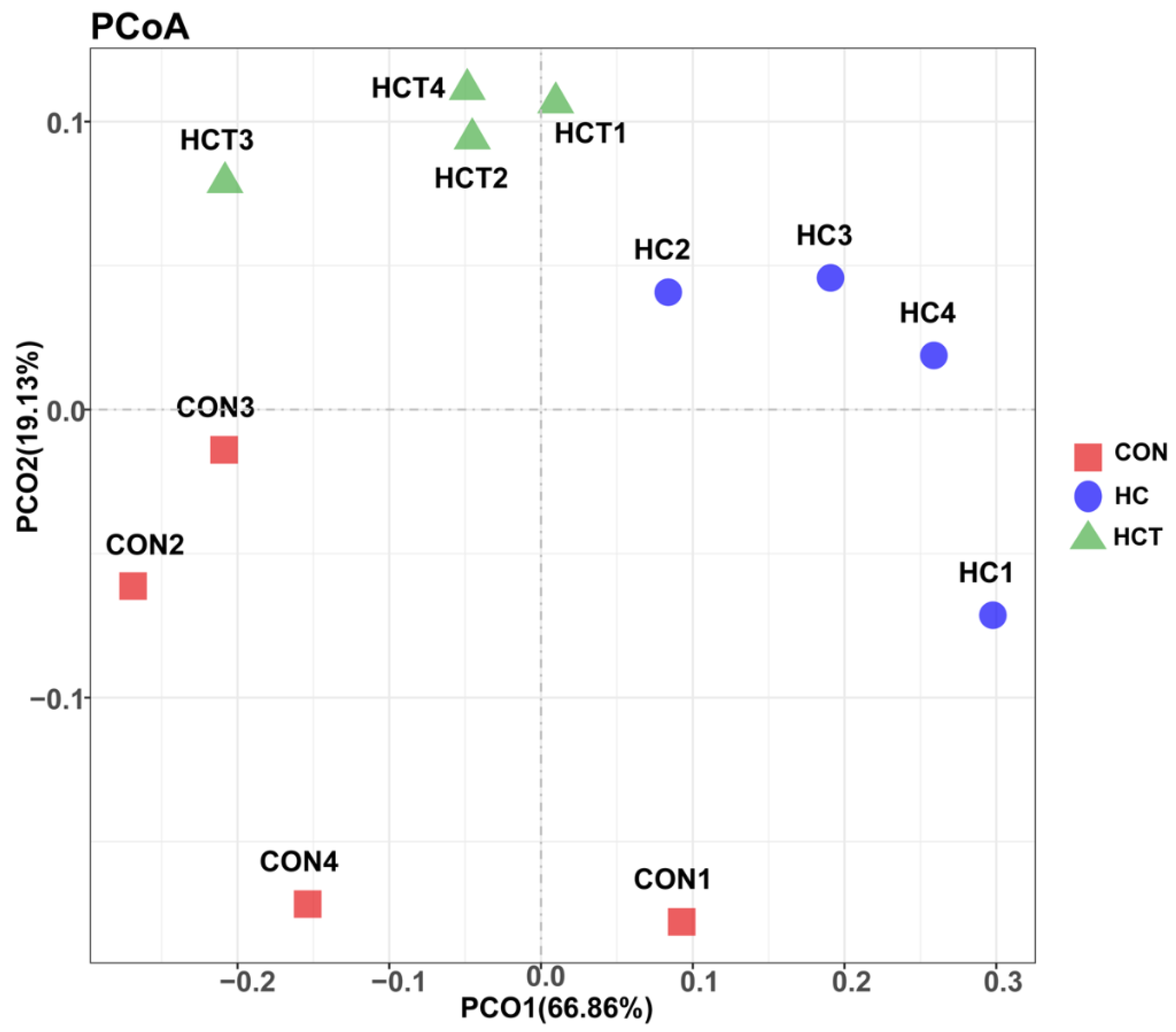
3.2. Taxonomy Results of Ruminal Bacteria and Methanongens
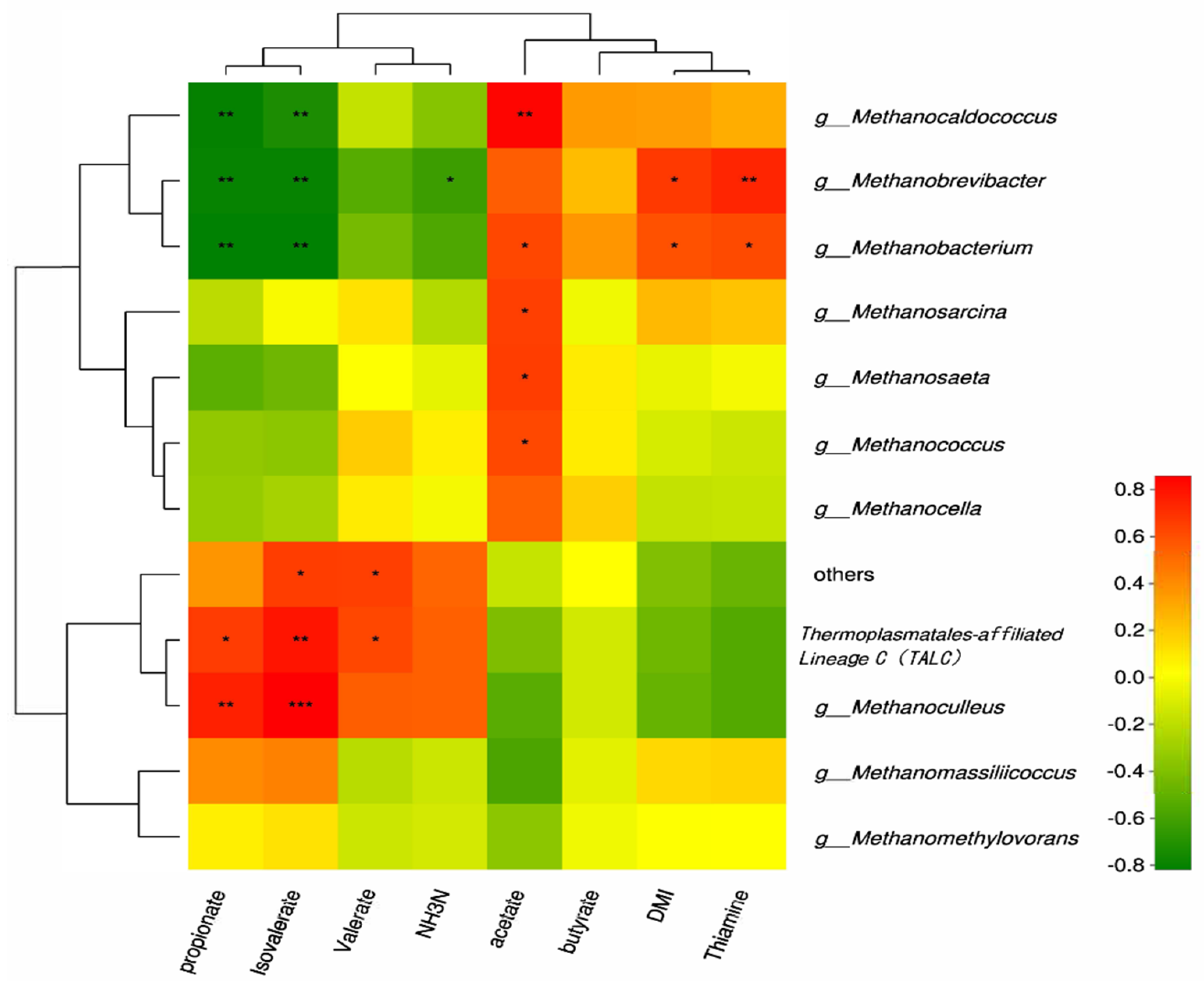
3.3. Effect of Thiamine Supplementation in High-Concentrate Diets on Ruminal Methanogenesis
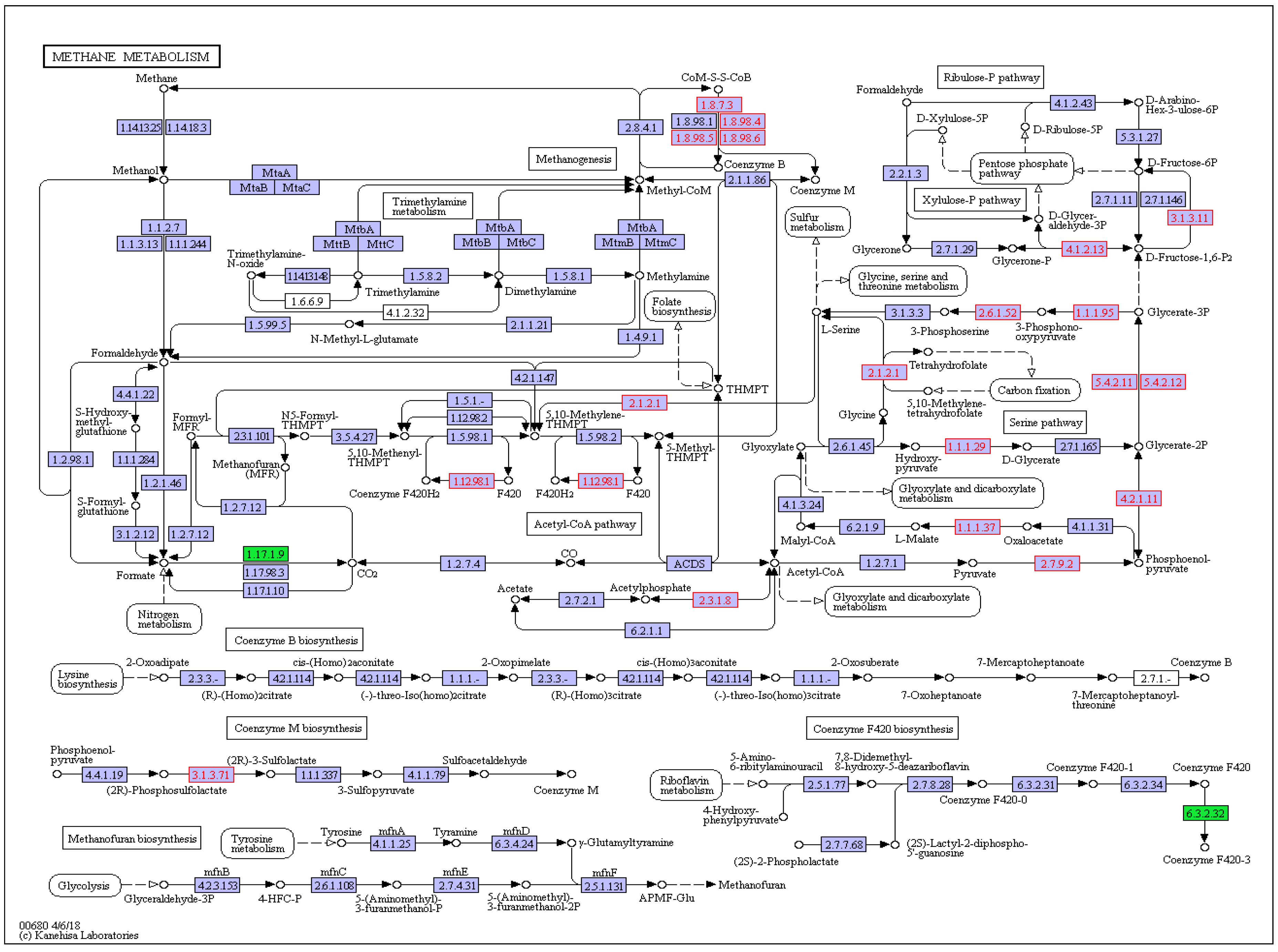
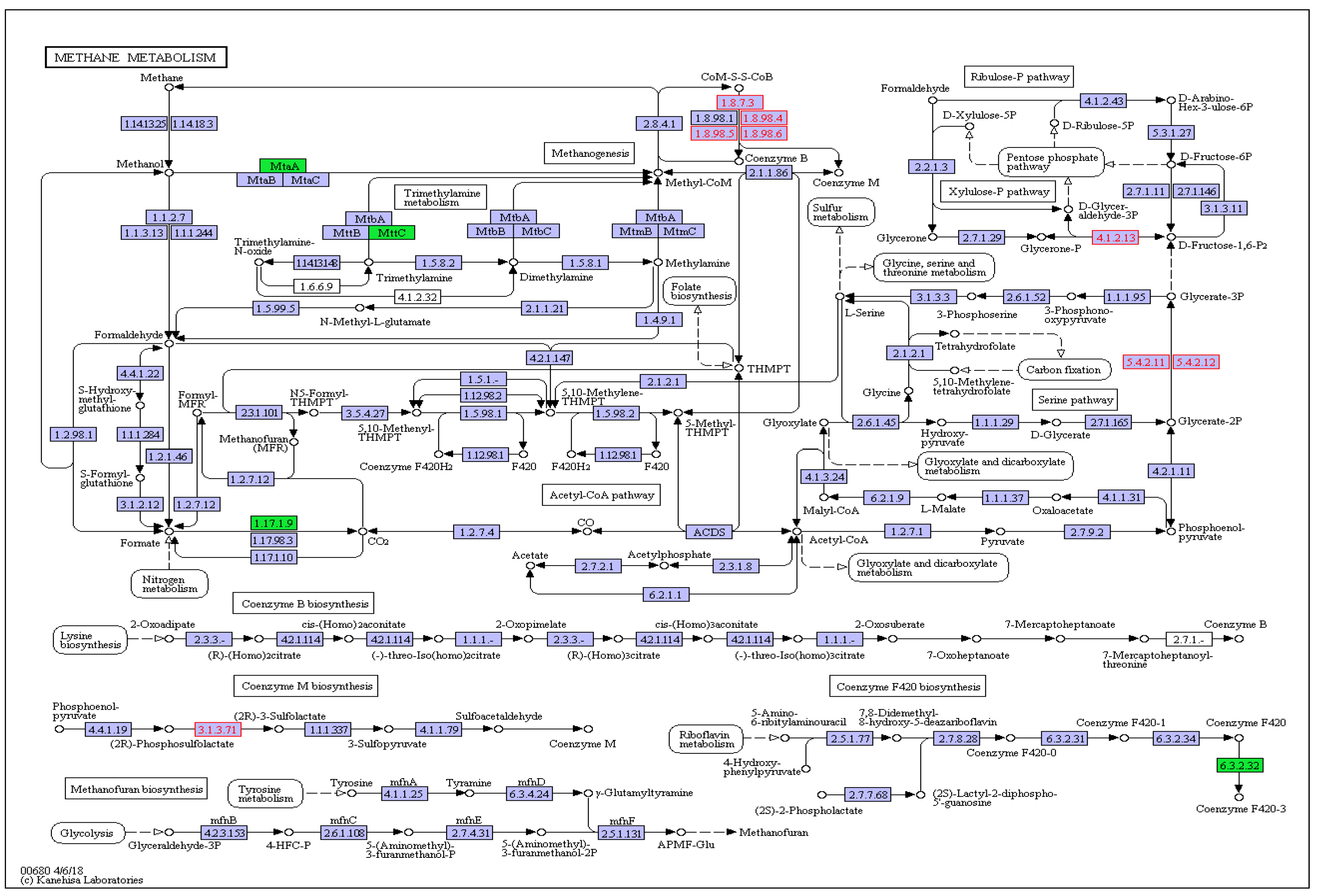
3.4. Functional Analysis of Thiamine Supplementation on Ruminal CH4 Metabolism
4. Discussion
4.1. Effects of Thiamine Supplementation in High-Concentrate Diets on Ruminal Methanogens
4.2. Effects of Thiamine Supplementation on CH4 Metabolism and CH4 Production
4.3. Comparison of Thiamine Supplementation in HC with Other CH4 Mitigation Strategies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hook, S.E.; Wright, A.G.; Mcbride, B.W. Methanogens: Methane Producers of the Rumen and Mitigation Strategies. Archaea Int. Microbiol. J. 2010, 2010, 945785. [Google Scholar] [CrossRef] [Green Version]
- Grainger, C.; Auldist, M.J.; Clarke, T.; Beauchemin, K.A.; Mcginn, S.M.; Hannah, M.C.; Eckard, R.J.; Lowe, L.B. Use of monensin controlled-release capsules to reduce methane emissions and improve milk production of dairy cows offered pasture supplemented with grain. J. Dairy Sci. 2008, 91, 1159–1165. [Google Scholar] [CrossRef]
- Hristov, A.N.; Oh, J.; Giallongo, F.; Frederick, T.W.; Harper, M.T.; Weeks, H.L.; Branco, A.F.; Moate, P.J.; Deighton, M.H.; Williams, S.R.; et al. An inhibitor persistently decreased enteric methane emission from dairy cows with no negative effect on milk production. Proc. Natl. Acad. Sci. USA 2015, 112, 10663–10668. [Google Scholar] [CrossRef] [Green Version]
- Adg, W.; Klieve, A.V. Does the complexity of the rumen microbial ecology preclude methane mitigation. 2011, 166–167, 248–253. Anim. Feed Sci. Technol. 2011, 166–167, 248–253. [Google Scholar]
- De Poorter, L.M.; Geerts, W.J.; Keltjens, J.T. Hydrogen concentrations in methane-forming cells probed by the ratios of reduced and oxidized coenzyme F420. Microbiology 2005, 151, 1697–1705. [Google Scholar] [CrossRef] [Green Version]
- Chellapandi, P.; Bharathi, M.; Sangavai, C.; Prathiviraj, R. Methanobacterium formicicum as a target rumen methanogen for the development of new methane mitigation interventions: A review. Vet. Anim. Sci. 2018, 6, 86–94. [Google Scholar] [CrossRef]
- Benchaar, C.; Hassanat, F.; Gervais, R.; Chouinard, P.Y.; Petit, H.V.; Masse, D.I. Methane production, digestion, ruminal fermentation, nitrogen balance, and milk production of cows fed corn silage- or barley silage-based diets. J. Dairy Sci. 2014, 97, 961–974. [Google Scholar] [CrossRef]
- Hall, M.B.; Mertens, D.R. A 100-Year Review: Carbohydrates-Characterization, digestion, and utilization. J. Dairy Sci. 2017, 100, 10078–10093. [Google Scholar] [CrossRef] [Green Version]
- Xue, F.; Pan, X.; Jiang, L.; Guo, Y.; Xiong, B. GC-MS analysis of the ruminal metabolome response to thiamine supplementation during high grain feeding in dairy cows. Metabolomics 2018, 14, 67. [Google Scholar] [CrossRef] [Green Version]
- Danielsson, R.; Dicksved, J.; Sun, L.; Gonda, H.; Muller, B.; Schnurer, A.; Bertilsson, J. Methane Production in Dairy Cows Correlates with Rumen Methanogenic and Bacterial Community Structure. Front. Microbiol. 2017, 8, 226. [Google Scholar] [CrossRef]
- Yu, Z.; García-González, R.; Schanbacher, F.L.; Morrison, M. Evaluations of Different Hypervariable Regions of Archaeal 16S rRNA Genes in Profiling of Methanogens by Archaea-Specific PCR and Denaturing Gradient Gel Electrophoresis. Appl. Environ. Microbiol. 2008, 74, 889–893. [Google Scholar] [CrossRef] [Green Version]
- Janssen, P.H.; Kirs, M. Structure of the archaeal community of the rumen. Appl. Environ. Microbiol. 2008, 74, 3619–3625. [Google Scholar] [CrossRef] [Green Version]
- Paul, K.; Nonoh, J.O.; Mikulski, L.; Brune, A. “Methanoplasmatales,” Thermoplasmatales-Related Archaea in Termite Guts and Other Environments, Are the Seventh Order of Methanogens. Appl. Environ. Microbiol. 2012, 78, 8245–8253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hess, M.; Sczyrba, A.; Egan, R.; Kim, T.W.; Chokhawala, H.; Schroth, G.; Luo, S.; Clark, D.S.; Chen, F.; Zhang, T. Metagenomic Discovery of Biomass-Degrading Genes and Genomes from Cow Rumen. Science 2011, 331, 463–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitta, D.W.; Indugu, N.; Kumar, S.; Vecchiarelli, B.; Sinha, R.; Baker, L.D.; Bhukya, B.; Ferguson, J.D. Metagenomic assessment of the functional potential of the rumen microbiome in Holstein dairy cows. Anaerobe 2016, 38, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Ranilla, M.J.; Carro, M.D.; López, S.; Newbold, C.J.; Wallace, R.J. Influence of nitrogen source on the fermentation of fibre from barley straw and sugarbeet pulp by ruminal micro-organisms in vitro. Br. J. Nutr. 2001, 86, 717. [Google Scholar] [CrossRef] [Green Version]
- Xue, F.; Nan, X.; Li, Y.; Pan, X.; Guo, Y.; Jiang, L.; Xiong, B. Metagenomic insights into effects of thiamine supplementation on ruminal non-methanogen archaea in high-concentrate diets feeding dairy cows. BMC Vet. Res. 2019, 15, 7. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, L.X.; Fang, S.X.; Zhang, F.; Liu, L.X. Study on the in-situ measurement of atmospheric CH4 and CO by GC-FID method at the Shangdianzi GAW regional station. Environ. Sci. 2012, 33, 8–12. [Google Scholar]
- Fussell, R.J.; Mccalley, D.V. Determination of volatile fatty acids (C2—C5) and lactic acid in silage by gas chromatography. Analyst 1987, 112, 1213–1216. [Google Scholar] [CrossRef]
- Terry, S.A.; Ribeiro, G.d.O.; Gruninger, R.J.; Hunerberg, M.; Ping, S.; Chaves, A.V.; Burlet, J.; Beauchemin, K.A.; McAllister, T.A. Effect of humic substances on rumen fermentation, nutrient digestibility, methane emissions, and rumen microbiota in beef heifers1. J. Anim. Sci. 2018, 96, 9. [Google Scholar] [CrossRef]
- Xue, F.; Nan, X.; Sun, F.; Pan, X.; Guo, Y.; Jiang, L.; Xiong, B. Metagenome sequencing to analyze the impacts of thiamine supplementation on ruminal fungi in dairy cows fed high-concentrate diets. AMB Express 2018, 8, 159. [Google Scholar] [CrossRef]
- Aronesty, E. Comparison of Sequencing Utility Programs. Open Bioinform. J. 2013, 7, 1–8. [Google Scholar] [CrossRef]
- Li, R.; Zhu, H.; Ruan, J.; Qian, W.; Fang, X.; Shi, Z.; Li, Y.; Li, S.; Shan, G.; Kristiansen, K.; et al. De novo assembly of human genomes with massively parallel short read sequencing. Genome Res. 2010, 20, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Bao, Z.; Okubo, T.; Kubota, K.; Kasahara, Y.; Tsurumaru, H.; Anda, M.; Ikeda, S.; Minamisawa, K. Metaproteomic Identification of Diazotrophic Methanotrophs and Their Localization in Root Tissues of Field-Grown Rice Plants. Appl. Environ. Microbiol. 2014, 80, 5043–5052. [Google Scholar] [CrossRef] [Green Version]
- Qin, N.; Yang, F.; Li, A.; Prifti, E.; Chen, Y.; Shao, L.; Guo, J.; Le Chatelier, E.; Yao, J.; Wu, L.; et al. Alterations of the human gut microbiome in liver cirrhosis. Nature 2014, 513, 59–64. [Google Scholar] [CrossRef]
- Huson, D.H.; Beier, S.; Flade, I.; Gorska, A.; El-Hadidi, M.; Mitra, S.; Ruscheweyh, H.J.; Tappu, R. MEGAN Community Edition—Interactive Exploration and Analysis of Large-Scale Microbiome Sequencing Data. PLoS Comput. Biol. 2016, 12, e1004957. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Kumar, S.; Lee, G.H.; Chang, D.H.; Rhee, M.S.; Yoon, M.H.; Kim, B.C. Methanobrevibacter boviskoreani sp. nov., isolated from the rumen of Korean native cattle. Int. J. Syst. Evol. Microbiol. 2013, 63, 4196–4201. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.H.; Yang, L.; Xue, F.G.; Xin, H.R.; Jiang, L.S.; Xiong, B.H.; Beckers, Y. Relationship between thiamine and subacute ruminal acidosis induced by a high-grain diet in dairy cows. J. Dairy Sci. 2016, 99, 8790–8801. [Google Scholar] [CrossRef] [Green Version]
- Joblin, K.N.; Campbell, G.P.; Richardson, A.J.; Stewart, C.S. Fermentation of barley straw by anaerobic rumen bacteria and fungi in axenic culture and in co-culture with methanogens. Lett. Appl. Microbiol. 2010, 9, 195–197. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Cavicchioli, R. Archaea: Molecular and Cellular Biology; ASM Press: Washington, DC, USA, 2007; pp. 151–152. [Google Scholar]
- Patra, A.K.; Yu, Z. Effects of Essential Oils on Methane Production and Fermentation by, and Abundance and Diversity of, Rumen Microbial Populations. Appl. Environ. Microbiol. 2012, 78, 4271–4280. [Google Scholar] [CrossRef] [Green Version]
- Pal, K.; Patra, A.K.; Sahoo, A.; Kumawat, P.K. Evaluation of several tropical tree leaves for methane production potential, degradability and rumen fermentation in vitro. Livest. Sci. 2015, 180, 98–105. [Google Scholar] [CrossRef]
- Patra, A.K. The effect of dietary fats on methane emissions, and its other effects on digestibility, rumen fermentation and lactation performance in cattle: A meta-analysis. Livest. Sci. 2013, 155, 244–254. [Google Scholar] [CrossRef]
- Reynolds, C.K.; Humphries, D.J.; Kirton, P.; Kindermann, M.; Duval, S.; Steinberg, W. Effects of 3-nitrooxypropanol on methane emission, digestion, and energy and nitrogen balance of lactating dairy cows. J. Dairy Sci. 2014, 97, 3777–3789. [Google Scholar] [CrossRef]
- Zijderveld, S.M.; Van Gerrits, W.J.J.; Dijkstra, J.; Newbold, J.R.; Hulshof, R.B.A.; Perdok, H.B. Persistency of methane mitigation by dietary nitrate supplementation in dairy cows. J. Dairy Sci. 2011, 94, 4028–4038. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Experimental Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | HC | HCT | |||
| Ruminal pH | 6.35 a | 5.88 c | 6.12 b | 0.094 | 0.016 |
| Thiamine content (ug/L) | 12.16 a | 8.51 b | 13.53 a | 1.933 | <0.001 |
| Acetate (mmol/L) | 44.24 a | 41.62 b | 44.07 a | 1.273 | 0.038 |
| Propionate (mmol/L) | 12.49 b | 13.75 a | 11.44 c | 0.632 | 0.027 |
| Propionate/Acetate | 0.282 b | 0.330 a | 0.259 b | 0.023 | 0.043 |
| Butyrate (mmol/L) | 10.77 | 10.35 | 10.82 | 0.137 | 0.356 |
| Ammonia-N (mg/100 mL) | 11.49 b | 13.96 a | 11.27 b | 1.711 | 0.006 |
| Total gas production (mL·g−1 DM) | 118.4 a | 105.2 b | 112.3 ab | 3.444 | 0.016 |
| Methane (mL·g−1 DM) | 6.91 a | 5.48 b | 5.82 b | 0.416 | 0.001 |
| Items | Experimental Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | HC | HCT | |||
| Bacteroidetes | 55.78 a | 31.51 c | 39.91 b | 3.274 | 0.001 |
| Firmicutes | 10.26 b | 15.57 a | 8.86 b | 0.935 | 0.001 |
| Bacteria_noname | 6.34 a | 3.32 b | 3.89 b | 0.459 | 0.002 |
| Proteobacteria | 5.06 b | 6.83 a | 3.25 a | 0.769 | 0.169 |
| Fibrobacteres | 0.99 a | 0.64 b | 0.88 a | 0.058 | 0.028 |
| Spirochaetes | 0.95 | 1.00 | 0.72 | 0.066 | 0.187 |
| Actinobacteria | 0.42 | 0.71 | 0.60 | 0.044 | 0.008 |
| Verrucomicrobia | 0.32 | 0.34 | 0.47 | 0.031 | 0.07 |
| Chlamydiae | 0.21 | 0.59 | 0.23 | 0.059 | 0.001 |
| Cyanobacteria | 0.19 | 0.14 | 0.54 | 0.056 | 0.001 |
| Planctomycetes | 0.10 | 0.08 | 0.16 | 0.012 | 0.024 |
| Tenericutes | 0.08 | 0.12 | 0.19 | 0.017 | 0.001 |
| Fusobacteria | 0.07 | 0.12 | 0.11 | 0.011 | 0.158 |
| others | 0.38 | 0.44 | 0.49 | 0.016 | 0.002 |
| Methanogens | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | HC | HCT | |||
| Methanobrevibacter | 0.1330 a | 0.0429 c | 0.0996 b | 0.0139 | 0.008 |
| Thermoplasmatales-affiliated Lineage C(TALC) | 0.0916 b | 0.1723 a | 0.0970 b | 0.0155 | 0.040 |
| Methanomassiliicoccus | 0.0376 | 0.0129 | 0.0068 | 0.0067 | 0.131 |
| Methanosarcina | 0.0191 | 0.0177 | 0.0241 | 0.0014 | 0.160 |
| Methanobacterium | 0.0099 a | 0.0032 b | 0.0089 a | 0.0012 | 0.036 |
| Methanoculleus | 0.0068 b | 0.0115 a | 0.0066 b | 0.0010 | 0.050 |
| Methanocaldococcus | 0.0031 b | 0.0017 c | 0.0062 a | 0.0006 | 0.001 |
| Methanococcus | 0.0015 b | 0.0021 b | 0.0064 a | 0.0007 | <0.001 |
| Methanosaeta | 0.0014 b | 0.0016 b | 0.0030 a | 0.0002 | 0.006 |
| Methanocella | 0.0014 b | 0.0018 b | 0.0063 a | 0.0007 | <0.001 |
| others | 0.0512 | 0.0740 | 0.0652 | 0.0046 | 0.123 |
| Total | 0.3581 | 0.3429 | 0.3304 | 0.1553 | 0.783 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, F.; Wang, Y.; Zhao, Y.; Nan, X.; Hua, D.; Sun, F.; Yang, L.; Jiang, L.; Xiong, B. Ruminal Methanogenic Responses to the Thiamine Supplementation in High-Concentrate Diets. Animals 2020, 10, 935. https://doi.org/10.3390/ani10060935
Xue F, Wang Y, Zhao Y, Nan X, Hua D, Sun F, Yang L, Jiang L, Xiong B. Ruminal Methanogenic Responses to the Thiamine Supplementation in High-Concentrate Diets. Animals. 2020; 10(6):935. https://doi.org/10.3390/ani10060935
Chicago/Turabian StyleXue, Fuguang, Yue Wang, Yiguang Zhao, Xuemei Nan, Dengke Hua, Fuyu Sun, Liang Yang, Linshu Jiang, and Benhai Xiong. 2020. "Ruminal Methanogenic Responses to the Thiamine Supplementation in High-Concentrate Diets" Animals 10, no. 6: 935. https://doi.org/10.3390/ani10060935
APA StyleXue, F., Wang, Y., Zhao, Y., Nan, X., Hua, D., Sun, F., Yang, L., Jiang, L., & Xiong, B. (2020). Ruminal Methanogenic Responses to the Thiamine Supplementation in High-Concentrate Diets. Animals, 10(6), 935. https://doi.org/10.3390/ani10060935