
Nigella sativa Extract Potentially Inhibited Methicillin Resistant Staphylococcus aureus Induced Infection in Rabbits: Potential Immunomodulatory and Growth Promoting Properties

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples Collection and Isolation and Characterization of MRSA
2.2. Preparation of NSE
2.3. In Vitro Antimicrobial Susceptibility Assays
2.4. Disc Diffusion Testing of MRSA Exposed to Sub-MICs of NSE
2.5. Synergism between Antibiotics and NSE
2.6. In Vivo Assessment of NSE Efficacy
2.6.1. Rabbits, Feeding and Experimental Design
2.6.2. Growth Performance
2.6.3. Sample Collection
2.6.4. Measurement of Digestive Enzymes and Serum Bactericidal Activities
2.6.5. Reverse Transcription Quantitative Real-Time PCR (qRT-PCR) Assays
2.6.6. Challenge Trial
2.7. Statistical Analysis
3. Results
3.1. Characterization and Incidence of MRSA Isolates
3.2. In Vitro Antimicrobial Susceptibility Patterns of MRSA Isolates
3.3. Antimicrobial Activity of NSE against MRSA Isolates
3.4. Antimicrobial Susceptibility Profiles of MRSA after Exposure to Sub-MICs of NSE
3.5. Synergism Assays between NSE and Antibiotics
3.6. Growth Performance and Digestive Enzymes’ Activities
3.7. Bactericidal Activity of Sera of NSE Fed Rabbits against MRSA Isolates
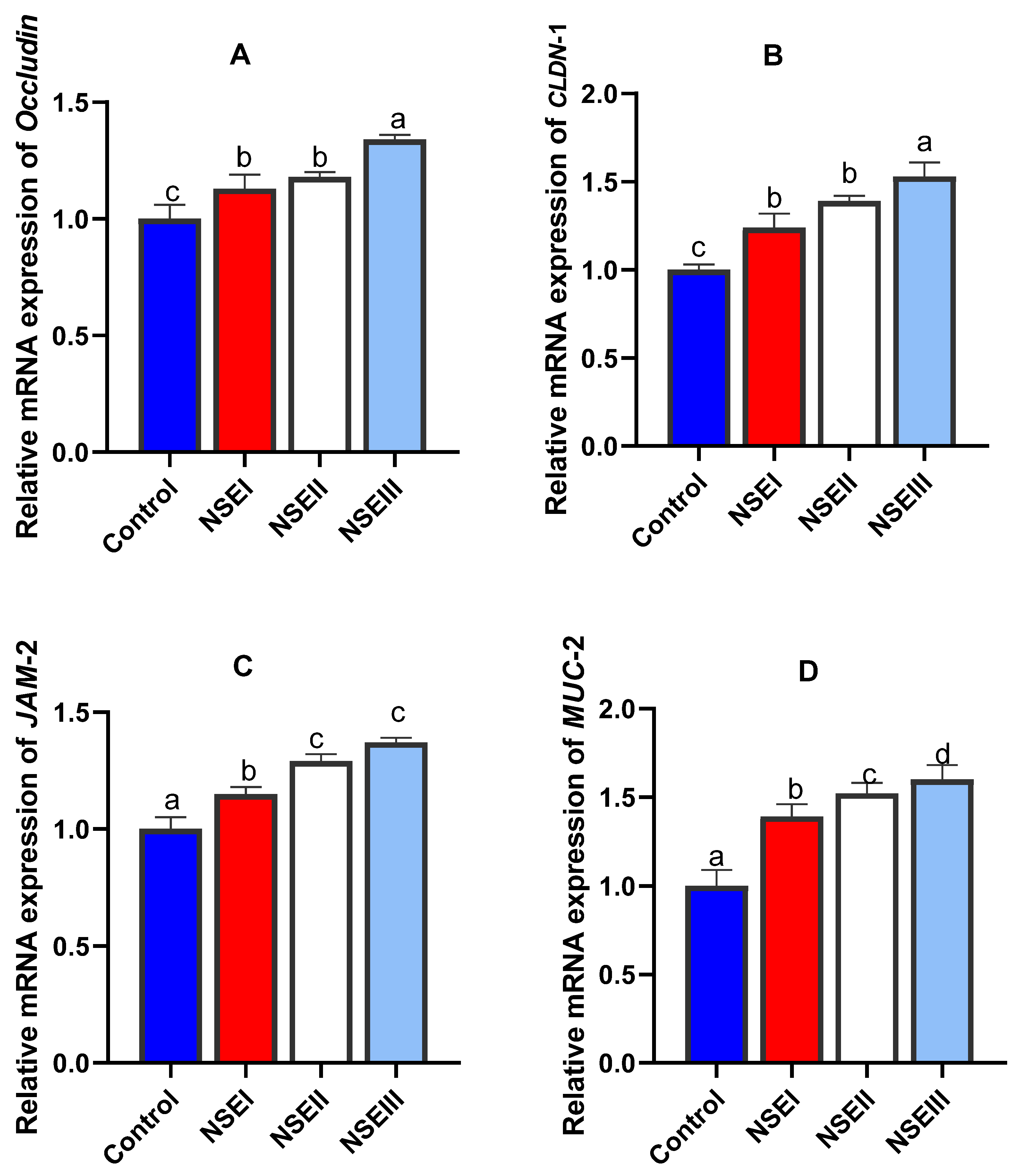
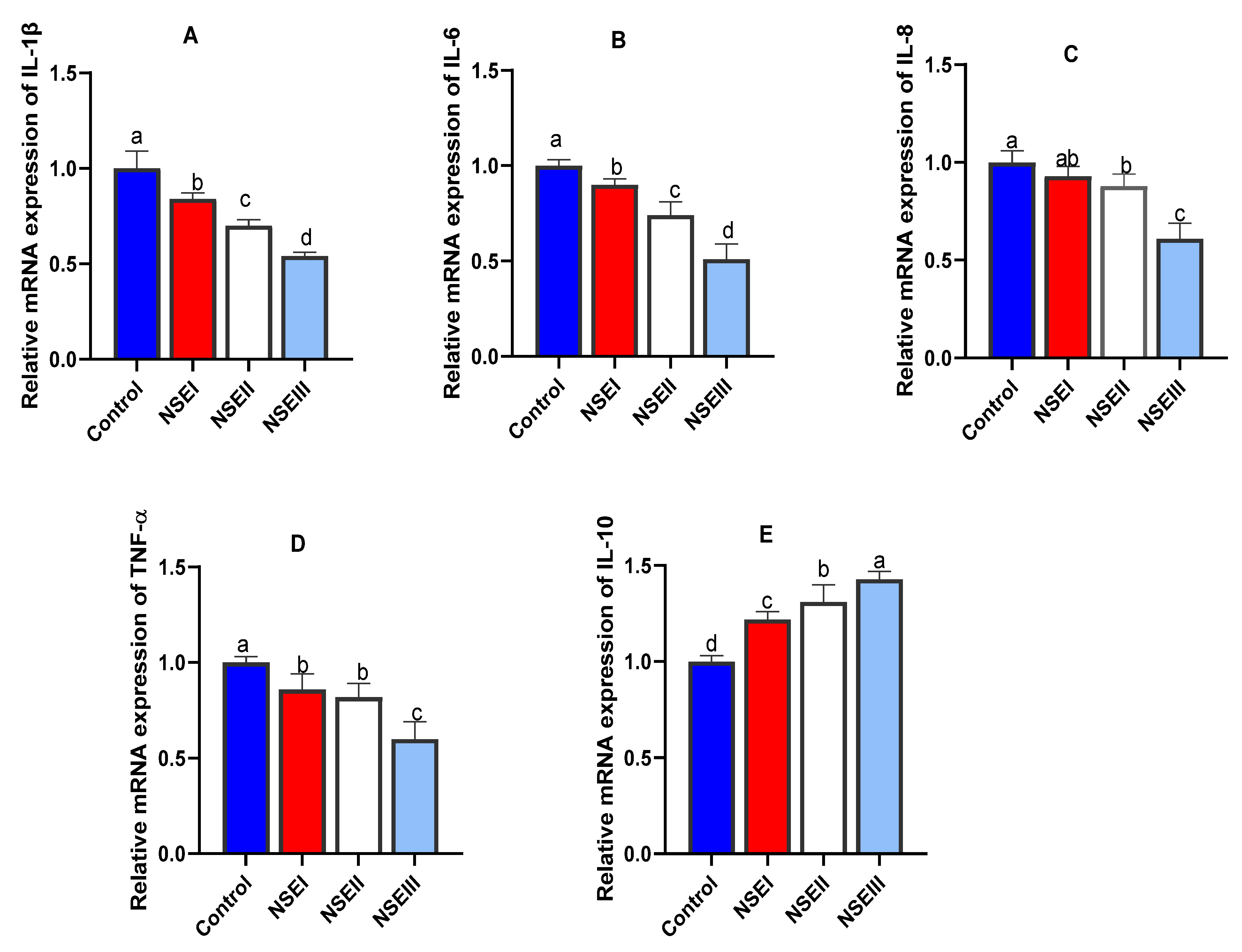
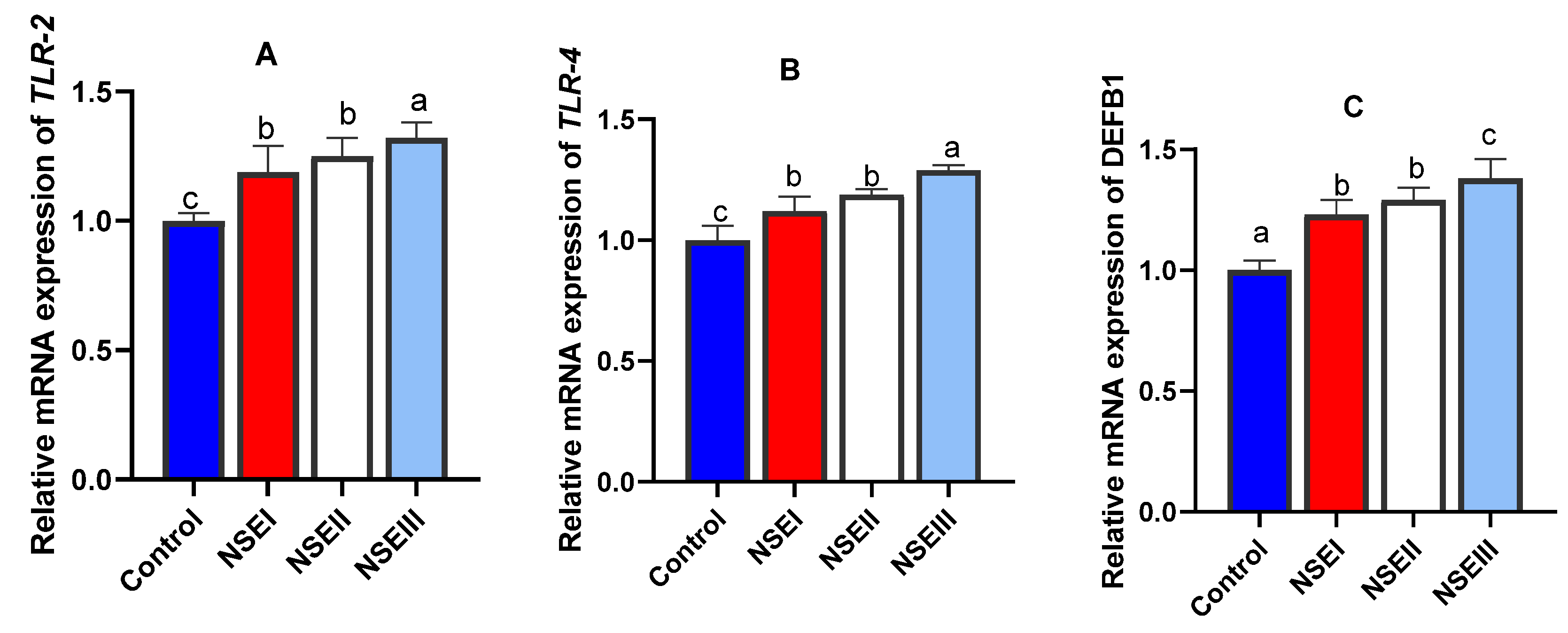
3.8. Gene Expression Analysis
3.9. Clearance of Rabbits’ Internal Organs from MRSA Infection by NSE
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christaki, E.; Marcou, M.; Tofarides, A. Antimicrobial resistance in bacteria: Mechanisms, evolution, and persistence. J. Mol. Evol. 2020, 88, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Girard, M.; Bee, G. Invited review: Tannins as a potential alternative to antibiotics to prevent coliform diarrhea in weaned pigs. Animal 2020, 14, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Alandiyjany, M.N.; Abdelaziz, A.S.; Abdelfattah-Hassan, A.; Hegazy, W.A.; Hassan, A.A.; Elazab, S.T.; Mohamed, E.A.; El-Shetry, E.S.; Saleh, A.A.; ElSawy, N.A. Novel In vivo assessment of antimicrobial efficacy of ciprofloxacin loaded lesoporous silica nanoparticles against Salmonella Typhimurium infection. Pharmaceuticals 2022, 15, 357. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, D.; Eldemery, F.; Metwally, A.S.; Abd-Allah, E.M.; Mohamed, D.T.; Ismail, T.A.; Hamed, T.A.; Al Sadik, G.M.; Neamat-Allah, A.N.; Abd El-Hamid, M.I. Dietary eugenol nanoemulsion potentiated performance of broiler chickens: Orchestration of digestive enzymes, intestinal barrier functions and cytokines related gene expression with a consequence of attenuating the severity of E. coli O78 infection. Front. Vet. Sci. 2022, 9, 847580. [Google Scholar] [CrossRef]
- Chambers, H.F.; DeLeo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef]
- Van Hal, S.J.; Jensen, S.O.; Vaska, V.L.; Espedido, B.A.; Paterson, D.L.; Gosbell, I.B. Predictors of mortality in Staphylococcus aureus bacteremia. Clin. Microbiol. Rev. 2012, 25, 362–386. [Google Scholar] [CrossRef]
- Gould, I.M.; David, M.Z.; Esposito, S.; Garau, J.; Lina, G.; Mazzei, T.; Peters, G. New insights into meticillin-resistant Staphylococcus aureus (MRSA) pathogenesis, treatment and resistance. Int. J. Antimicrob. Agents 2012, 39, 96–104. [Google Scholar] [CrossRef]
- Ammar, A.; El-Hamid, M.; Eid, S.E.; Oksh, A.S.E. Insight into antimicrobial resistance and virulence genes of emergent multidrug resistant avian pathogenic Escherichia coli in Egypt: How closely related are they. Rev. Med. Vet. 2015, 166, 304–314. [Google Scholar]
- Ammar, A.M.; El-Naenaeey, E.-S.Y.; El-Hamid, M.I.A.; El-Gedawy, A.A.; Elmalt, R.M. Campylobacter as a major foodborne pathogen: A review of its characteristics, pathogenesis, antimicrobial resistance and control. J. Microbiol. Biotechnol. Food Sci. 2021, 10, 609–619. [Google Scholar] [CrossRef]
- Hashem, Y.M.; El-Hamid, M.I.A.; Awad, N.F.; Ibrahim, D.; Elshater, N.S.; El-Malt, R.M.; Hassan, W.H.; Abo-Shama, U.H.; Nassan, M.A.; El-Bahy, S.M. Insights into growth-promoting, anti-inflammatory, immunostimulant, and antibacterial activities of Toldin CRD as a novel phytobiotic in broiler chickens experimentally infected with Mycoplasma gallisepticum. Poult. Sci. 2022, 101, 102154. [Google Scholar] [CrossRef]
- El-Hamid, M.I.A.; Ibrahim, D.; Hamed, R.I.; Nossieur, H.H.; Elbanna, M.H.; Baz, H.; Abd-Allah, E.M.; Oksh, A.S.E.; Ibrahim, G.A.; Khalifa, E. Modulatory impacts of multi-strain probiotics on rabbits’ growth, nutrient transporters, tight junctions and immune system to fight against Listeria monocytogenes infection. Animals 2022, 12, 2082. [Google Scholar] [CrossRef] [PubMed]
- Khater, S.I.; Lotfy, M.M.; Alandiyjany, M.N.; Alqahtani, L.S.; Zaglool, A.W.; Althobaiti, F.; Ismail, T.A.; Soliman, M.M.; Saad, S.; Ibrahim, D. Therapeutic potential of quercetin loaded nanoparticles: Novel insights in alleviating colitis in an experimental DSS induced colitis model. Biomedicines 2022, 10, 1654. [Google Scholar] [CrossRef] [PubMed]
- Aljazzar, A.; El-Hamid, M.I.A.; El-Malt, R.M.; El-Gharreb, W.R.; Abdel-Raheem, S.M.; Ibrahim, A.M.; Abdelaziz, A.M.; Ibrahim, D. Prevalence and antimicrobial susceptibility of Campylobacter species with particular focus on the growth promoting, immunostimulant and anti-Campylobacter jejuni activities of eugenol and trans-cinnamaldehyde mixture in broiler chickens. Animals 2022, 12, 905. [Google Scholar] [CrossRef]
- Ibrahim, D.; Moustafa, A.; Metwally, A.S.; Nassan, M.A.; Abdallah, K.; Eldemery, F.; Tufarelli, V.; Laudadio, V.; Kishawy, A.T. Potential application of cornelian cherry extract on broiler chickens: Growth, expression of antioxidant biomarker and glucose transport genes, and oxidative stability of frozen meat. Animals 2021, 11, 1038. [Google Scholar] [CrossRef] [PubMed]
- Ammar, A.; Attia, A.; El-Hamid, M.A.; El-Shorbagy, I.; El-Kader, S.A. Genetic basis of resistance waves among methicillin resistant Staphylococcus aureus isolates recovered from milk and meat products in Egypt. Cell. Mol. Biol. 2016, 62, 7–15. [Google Scholar]
- Toama, M.A.; El-Alfy, T.S.; El-Fatatry, H.M. Antimicrobial activity of the volatile oil of Nigella sativa Linneaus seeds. Antimicrob. Agents Chemother. 1974, 6, 225–226. [Google Scholar] [CrossRef] [PubMed]
- Kumar, T.S.; Negi, P.; Sankar, K.U. Antibacterial activity of “Nigella sativa L.” seed extracts. Br. J. Pharmacol. Toxicol. 2010, 1, 96–100. [Google Scholar]
- Chaieb, K.; Kouidhi, B.; Jrah, H.; Mahdouani, K.; Bakhrouf, A. Antibacterial activity of thymoquinone, an active principle of Nigella sativa and its potency to prevent bacterial biofilm formation. BMC Complement. Altern. Med. 2011, 11, 29. [Google Scholar] [CrossRef]
- Abu-Al-Basalc, M.A. In vitro and iin vivo anti-microbial effects of Nigella sativa Linn. seed extracts against clinical isolates from skin wound infections. Am. J. Appl. Sci. 2009, 6, 1440. [Google Scholar]
- Sharikh, S.M.; Rao, J.U.P.; Ackshaya, M.S. A study of antibacterial effect of Nigella Sativa seed extract on clinical isolates of methicillin-resistant Staphylococcus aureus (MRSA). Indian J. Public Health Res. Dev. 2020, 11, 1516–1520. [Google Scholar]
- Nagi, A.; Mariana, N.; Hana, F.; Rasedee, A. Extract of essential iol from Nigella sativa using superfacial carbon dioxide: Study of antibacterial activity. Am. J. Pharmacol. Toxicol. 2008, 3, 225–228. [Google Scholar]
- El-Hamid, M.I.A.; El-Sayed, M.; Ali, A.R.; Abdallah, H.; Arnaout, M.I.; El-Mowalid, G.A. Marjoram extract down-regulates the expression of Pasteurella multocida adhesion, colonization and toxin genes: A potential mechanism for its antimicrobial activity. Comp. Immunol. Microbiol. Infect. Dis. 2019, 62, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Piras, A.; Rosa, A.; Marongiu, B.; Porcedda, S.; Falconieri, D.; Dessi, M.A.; Ozcelik, B.; Koca, U. Chemical composition and iin vitro bioactivity of the volatile and fixed oils of Nigella sativa L. extracted by supercritical carbon dioxide. Ind. Crops Prod. 2013, 46, 317–323. [Google Scholar] [CrossRef]
- Halawani, E. Antibacterial activity of thymoquinone and thymohydroquinone of Nigella sativa L. and their interaction with some antibiotics. Adv. Biol. Res. 2009, 3, 148–152. [Google Scholar]
- El-Hamid, M.A.; Bendary, M.; Merwad, A.; Elsohaby, I.; Mohammad Ghaith, D.; Alshareef, W. What is behind phylogenetic analysis of hospital-, community-and livestock-associated methicillin-resistant Staphylococcus aureus? Transbound. Emerg. Dis. 2019, 66, 1506–1517. [Google Scholar]
- El-Hamid, A.; Marwa, I.; Sewid, A.H.; Samir, M.; Hegazy, W.A.; Bahnass, M.M.; Mosbah, R.A.; Ghaith, D.M.; Khalifa, E.; Ramadan, H. Clonal diversity and epidemiological characteristics of ST239-MRSA Strains. Front. Cell. Infect. Microbiol. 2022, 12, 782045. [Google Scholar] [CrossRef]
- El-Hamid, M.I.A.; Bendary, M. Comparative phenotypic and genotypic discrimination of methicillin resistant and susceptible Staphylococcus aureus in Egypt. Cell. Mol. Biol. 2015, 61, 101–112. [Google Scholar]
- El-Aziz, N.K.A.; El-Hamid, M.I.A.; Bendary, M.M.; El-Azazy, A.A.; Ammar, A.M. Existence of vancomycin resistance among methicillin resistant S. aureus recovered from animal and human sources in Egypt. Slov. Vet. Res. 2018, 55, 221–230. [Google Scholar]
- Hanafy, M.; Hatem, M. Studies on the antimicrobial activity of Nigella sativa seed (black cumin). J. Ethnopharmacol. 1991, 34, 275–278. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute Performance standards for Antimicrobial Susceptibility Testing; Twenty-First Informational Supplement. CLSI Document M100-S21. Clinical and Laboratory Standards Institute, Wayne. Available online: http://www.antimicrobianos.com.ar/ATB/wpcontent/M100S21E.pdf (accessed on 1 January 2011).
- Ahmed, H.A.; Tahoun, A.B.; Elez, R.M.A.; El-Hamid, M.I.A.; Ellatif, S.S.A. Prevalence of Yersinia enterocolitica in milk and dairy products and the effects of storage temperatures on survival and virulence gene expression. Int. Dairy J. 2019, 94, 16–21. [Google Scholar] [CrossRef]
- Reller, L.B.; Stratton, C.W. Serum dilution test for bactericidal activity. II. Standardization and correlation with antimicrobial assays and susceptibility tests. J. Infect. Dis. 1977, 136, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Blas, C.D.; Wiseman, J. CAB International, Wallingford Oxon (UK); The Nutrition of the rabbit; CAB Publishing, CAB International: Wallingford, Oxon UK, 1998; p. 3441. [Google Scholar]
- Ibrahim, D.; Abdelfattah-Hassan, A.; Arisha, A.H.; El-Aziz, R.M.A.; Sherief, W.R.; Adil, S.H.; El Sayed, R.; Metwally, A.E. Impact of feeding anaerobically fermented feed supplemented with acidifiers on its quality and growth performance, intestinal villi and enteric pathogens of mulard ducks. Livest. Sci. 2020, 242, 104299. [Google Scholar] [CrossRef]
- Ibrahim, D.; Sewid, A.H.; Arisha, A.H.; El-Fattah, A.H.A.; Abdelaziz, A.M.; Al-Jabr, O.A.; Kishawy, A.T. Influence of glycyrrhiza glabra extract on growth, gene expression of gut integrity, and Campylobacter jejuni colonization in broiler chickens. Front. Vet. Sci. 2020, 7, 612063. [Google Scholar] [CrossRef] [PubMed]
- Farahat, M.; Ibrahim, D.; Kishawy, A.; Abdallah, H.; Hernandez-Santana, A.; Attia, G. Effect of cereal type and plant extract addition on the growth performance, intestinal morphology, caecal microflora, and gut barriers gene expression of broiler chickens. Animal 2020, 15, 100056. [Google Scholar] [CrossRef]
- Omar, A.E.; Al-Khalaifah, H.S.; Ismail, T.A.; El-Aziz, R.M.A.; El-Mandrawy, S.A.; Shalaby, S.I.; Ibrahim, D. Performance, serum biochemical and immunological parameters, and digestive enzyme and intestinal barrier-related gene expression of broiler chickens fed fermented fava bean by-products as a substitute for conventional feed. Front. Vet. Sci. 2021, 8, 696841. [Google Scholar] [CrossRef]
- Blasco, A.; Ouhayoun, J. Harmonization of criteria and terminology in rabbit meat research. Revised proposal. World Rabbit Sci. 1996, 4, 93–99. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ibrahim, D.; Ismail, T.A.; Khalifa, E.; El-Kader, A.; Shaimaa, A.; Mohamed, D.I.; Mohamed, D.T.; Shahin, S.E.; El-Hamid, A.; Marwa, I. Supplementing garlic nanohydrogel optimized growth, gastrointestinal integrity and economics and ameliorated necrotic enteritis in broiler chickens using a Clostridium perfringens challenge model. Animals 2021, 11, 2027. [Google Scholar] [CrossRef]
- El-Hamid, M.I.A.; Ibrahim, S.M.; Eldemery, F.; El-Mandrawy, S.A.; Metwally, A.S.; Khalifa, E.; Elnahriry, S.S.; Ibrahim, D. Dietary cinnamaldehyde nanoemulsion boosts growth and transcriptomes of antioxidant and immune related genes to fight Streptococcus agalactiae infection in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2021, 113, 96–105. [Google Scholar] [CrossRef]
- Ibrahim, D.; Kishawy, A.T.; Khater, S.I.; Khalifa, E.; Ismail, T.A.; Mohammed, H.A.; Elnahriry, S.S.; Tolba, H.A.; Sherief, W.R.; Farag, M.F. Interactive effects of dietary quercetin nanoparticles on growth, flesh antioxidant capacity and transcription of cytokines and Aeromonas hydrophila quorum sensing orchestrating genes in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2021, 119, 478–489. [Google Scholar] [CrossRef]
- Ahmad, A.; Husain, A.; Mujeeb, M.; Khan, S.A.; Najmi, A.K.; Siddique, N.A.; Damanhouri, Z.A.; Anwar, F. A review on therapeutic potential of Nigella sativa: A miracle herb. Asian Pac. J. Trop. Biomed. 2013, 3, 337–352. [Google Scholar] [CrossRef]
- Hannan, A.; Saleem, S.; Chaudhary, S.; Barkaat, M.; Arshad, M.U. Anti bacterial activity of Nigella sativa against clinical isolates of methicillin resistant Staphylococcus aureus. J. Ayub Med. Coll. Abbottabad 2008, 20, 72–74. [Google Scholar] [PubMed]
- Hartmann, M.; Berditsch, M.; Hawecker, J.; Ardakani, M.F.; Gerthsen, D.; Ulrich, A.S. Damage of the bacterial cell envelope by antimicrobial peptides gramicidin S and PGLa as revealed by transmission and scanning electron microscopy. Antimicrob. Agents Chemother. 2010, 54, 3132–3142. [Google Scholar] [CrossRef] [PubMed]
- Toghyani, M.; Toghyani, M.; Gheisari, A.; Ghalamkari, G.; Mohammadrezaei, M. Growth performance, serum biochemistry and blood hematology of broiler chicks fed different levels of black seed (Nigella sativa) and peppermint (Mentha piperita). Livest. Sci. 2010, 129, 173–178. [Google Scholar] [CrossRef]
- Jamroz, D.; Kamel, C. Plant extracts enhance broiler performance. In non-ruminant nutrition: Antimicrobial agents and plant extracts on immunity, health and performance. J. Anim. Sci. 2002, 80, 41. [Google Scholar]
- Kishawy, A.T.; Al-Khalaifah, H.S.; Nada, H.S.; Roushdy, E.M.; Zaglool, A.W.; Ahmed Ismail, T.; Ibrahim, S.M.; Ibrahim, D. Black pepper or radish seed oils in a new combination of essential oils modulated broiler chickens’ performance and expression of digestive enzymes, lipogenesis, immunity, and autophagy-related genes. Vet. Sci. 2022, 9, 43. [Google Scholar] [CrossRef]
- Al-Beitawi, N.; El-Ghousein, S. Effect of feeding different levels of Nigella sativa seeds (black cumin) on performance, blood constituents and carcass characteristics of broiler chicks. Int. J. Poult. Sci. 2008, 7, 715–721. [Google Scholar] [CrossRef]
- Hannan, M.A.; Rahman, M.A.; Sohag, A.A.M.; Uddin, M.J.; Dash, R.; Sikder, M.H.; Rahman, M.S.; Timalsina, B.; Munni, Y.A.; Sarker, P.P. Black cumin (Nigella sativa L.): A comprehensive review on phytochemistry, health benefits, molecular pharmacology, and safety. Nutrients 2021, 13, 1784. [Google Scholar] [CrossRef]
- Asghar, M.U.; Doğan, S.C.; Wilk, M.; Korczyński, M. Effect of dietary supplementation of black cumin seeds (Nigella sativa) on performance, carcass traits, and meat quality of Japanese quails (Coturnix coturnix japonica). Animals 2022, 12, 1298. [Google Scholar] [CrossRef]
- Tabler, T.W.; Greene, E.S.; Orlowski, S.K.; Hiltz, J.Z.; Anthony, N.B.; Dridi, S. Intestinal barrier integrity in heat-stressed modern broilers and their ancestor wild jungle fowl. Front. Vet. Sci. 2020, 7, 249. [Google Scholar] [CrossRef]
- Ibrahim, D.; Abdelfattah-Hassan, A.; Badawi, M.; Ismail, T.A.; Bendary, M.M.; Abdelaziz, A.M.; Mosbah, R.A.; Mohamed, D.I.; Arisha, A.H.; El-Hamid, M.I.A. Thymol nanoemulsion promoted broiler chicken’s growth, gastrointestinal barrier and bacterial community and conferred protection against Salmonella Typhimurium. Sci. Rep. 2021, 11, 7742. [Google Scholar] [CrossRef] [PubMed]
- Kapan, M.; Tekin, R.; Onder, A.; Firat, U.; Evliyaoglu, O.; Taskesen, F.; Arikanoglu, Z. Thymoquinone ameliorates bacterial translocation and inflammatory response in rats with intestinal obstruction. Int. J. Surg. 2012, 10, 484–488. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K. Influence of plant bioactive compounds on intestinal epithelial barrier in poultry. Mini Rev. Med. Chem. 2020, 20, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Amasheh, S.; Aschenbach, J.R. Modulation of gastrointestinal barrier and nutrient transport function in farm animals by natural plant bioactive compounds–a comprehensive review. Crit. Rev. Food Sci. Nutr. 2019, 59, 3237–3266. [Google Scholar] [CrossRef]
- Salem, M.L. Immunomodulatory and therapeutic properties of the Nigella sativa L. seed. Int. Immunopharmacol. 2005, 5, 1749–1770. [Google Scholar] [CrossRef]
- Al-Mufarrej, S. Immune-responsiveness and performance of broiler chickens fed black cumin (Nigella sativa L.) powder. J. Saudi Soc. Agric. Sci. 2014, 13, 75–80. [Google Scholar] [CrossRef]
- El-Bagir, N.M.; Farah, I.T.; Elhag, S.M.; Alhaidary, A.; Mohamed, H.E.; Beynen, A.C. Immune response and pasteurella resistance in rabbits fed diets containing various amounts of black cumin seeds. Am. J. Anim. Vet. Sci. 2010, 5, 163–167. [Google Scholar] [CrossRef]
- Mahmoud, H.S.; Almallah, A.A.; EL-Hak, H.N.G.; Aldayel, T.S.; Abdelrazek, H.; Khaled, H.E. The effect of dietary supplementation with Nigella sativa (black seeds) mediates immunological function in male Wistar rats. Sci. Rep. 2021, 11, 7542. [Google Scholar] [CrossRef]
- Amin, B.; Hosseinzadeh, H. Black cumin (Nigella sativa) and its active constituent, thymoquinone: An overview on the analgesic and anti-inflammatory effects. Planta Med. 2016, 82, 8–16. [Google Scholar] [CrossRef]
- Zaoui, A.; Cherrah, Y.; Alaoui, K.; Mahassine, N.; Amarouch, H.; Hassar, M. Effects of Nigella sativa fixed oil on blood homeostasis in rat. J. Ethnopharmacol. 2002, 79, 23–26. [Google Scholar] [CrossRef]
- Fararh, K.; Atoji, Y.; Shimizu, Y.; Shiina, T.; Nikami, H.; Takewaki, T. Mechanisms of the hypoglycaemic and immunopotentiating effects of Nigella sativa L. oil in streptozotocin-induced diabetic hamsters. Res. Vet. Sci. 2004, 77, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Haq, A.; Lobo, P.I.; Al-Tufail, M.; Rama, N.R.; Al-Sedairy, S.T. Immunomodulatory effect of Nigella sativa proteins fractionated by ion exchange chromatography. Int. J. Immunopharmacol. 1999, 21, 283–295. [Google Scholar] [CrossRef]
- Elmowalid, G.; Amar, A.M.; Ahmad, A.A.M. Nigella sativa seed extract: 1. Enhancement of sheep macrophage immune functions in vitro. Res. Vet. Sci. 2013, 95, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Majdalawieh, A.F.; Hmaidan, R.; Carr, R.I. Nigella sativa modulates splenocyte proliferation, Th1/Th2 cytokine profile, macrophage function and NK anti-tumor activity. J. Ethnopharmacol. 2010, 131, 268–275. [Google Scholar] [CrossRef]
- Salem, M.L.; Hossain, M.S. Protective effect of black seed oil from Nigella sativa against murine cytomegalovirus infection. Int. J. Immunopharmacol. 2000, 22, 729–740. [Google Scholar] [CrossRef]
- Boskabady, M.-H.; Keyhanmanesh, R.; Khameneh, S.; Doostdar, Y.; Khakzad, M.-R. Potential immunomodulation effect of the extract of Nigella sativa on ovalbumin sensitized guinea pigs. J. Zhejiang Univ. Sci. B 2011, 12, 201–209. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Ingredient | % |
|---|---|
| Yellow corn | 9.50 |
| Barley grains | 18.60 |
| Soybean meal, 44% | 16.50 |
| Berseem hay | 34.00 |
| Wheat bran | 15.80 |
| Molasses | 2.80 |
| Premix * | 0.3 |
| Calcium dibasic phosphate | 1.5 |
| Common salt | 0.5 |
| Antitoxin | 0.3 |
| Anticoccidial | 0.2 |
| Nutrient levels | |
| Digestable energy, Kcal/Kg | 2587 |
| Crude protien | 16.55 |
| Ether extract | 2.26 |
| Crude fiber | 12.60 |
| Ca | 1.02 |
| Phosphorus | 0.62 |
| Lysin | 0.76 |
| Methionine | 0.25 |
| Specificity/Encoding Gene | Primer Sequence (5′-3′) | Accession No |
|---|---|---|
| Tight junction proteins | ||
| MUC-2 | F: TATACCGCAAGCAGCCAGGT R: GCAAGCAGGACACAGACCAG | L41544.1 |
| JAM-2 | F: ATATCGCAGGTGTCCTGGAA R: GAGCATAGCACACGCCAAG | XM_017346699 |
| CLDN-1 | F: GGAGCAAAAGATGCGGATGG R: AATTGACAGGGGTCAAAGGGT | NM_001089316.1 |
| Occludin | F: GCAAGAGGCCGTATCCAGAG R: AGTCCGTCTCGTAGTGGTCT | XM_008262320.1 |
| Cytokines | ||
| IL-6 | F: GCCAACCCTACAACAAGA R: AGAGCCACAACGACTGAC | NC_013678 |
| IL-8 | F: CTCTCTTGGCAACCTTCCTG R: TTGCACAGTGAGGTCCACTC | KT216053.1 |
| IL-10 | F: AAAAGCTAAAAGCCCCAGGA R: CGGGAGCTGAGGTATCAGAG | NM001082045.1 |
| IL-1β | F: TTCCGGATGTATCTCGAGCA R: GTGGATCGTGGTCGTCTTCA | NC_013670 |
| TNF-α | F: CTGCACTTCAGGGTGATCG R: CTACGTGGGCTAGAGGCTTG | XM_008262537.2 |
| TLR-2 | F: TGCCTCCTTGTTACCTATGC R: AGATGAAGTTGTTCCCTCCG | NM_00108271 |
| TLR-4 | F: AGATGAAGTTGTTCCCTCCG R: GTGGGCTTAGAACAACTGGAAC | NM_001082732.2 |
| DEFB1 | F: AGCCTGTCTGCCTGGAGTAG R: GATGAGGAGAGGCTTCATGG | XM017337690.1 |
| House keeping | ||
| GAPDH | F: TGTTTGTGATGGGCGTGAA R: CCTCCACAATGCCGAAGT | NC_013676.1 |
| Sample Source | MRSA Tested Isolates | Antimicrobial Resistance Profile | NSE Antimicrobial Activity | |
|---|---|---|---|---|
| Zone Diameter (mm) | MIC (ug/mL) | |||
| Cows’ milk | 1 M | ME, E, DO, DA, AMC | 28 | 16 |
| 2 M | ME, AMC | 18 | 128 | |
| 3 M | ME, AMC | 30 | 8 | |
| 4 M | ME, SXT, CIP | - (R) | 1024 | |
| 5 M | ME | 12 (R) | 512 | |
| Meat products | 1 Mm | ME, AMC, CIP | 18 | 128 |
| 2 Br | ME, DO, CIP | 18 | 128 | |
| 3 Br | ME, AMC, DO, RF | 18 | 128 | |
| 5 Sg | ME, RF, E, DA, AMC | 15 | 256 | |
| Sample Source | MRSA Tested Isolates | Antimicrobial Resistance Profile | NSE Antimicrobial Activity | |
|---|---|---|---|---|
| Zone Diameter (mm) | MIC (ug/mL) | |||
| Pus | 1 Pu | ME, RF, DO, CN, SXT, AMC, E | 15 | 256 |
| 2 Pu | ME, SXT, AMC, DA, RF | 15 | 256 | |
| 3 Pu | ME, DO, SXT, DA, CIP | - (R) | 1024 | |
| 4 Pu | ME | 12 (R) | 512 | |
| 5 Pu | ME | 20 | 64 | |
| Urine | 1 U | ME, E, RF, AMC, SXT | 12 (R) | 512 |
| 2 U | ME, E, SXT | 12 (R) | 512 | |
| 3 U | ME, E, SXT, CIP | 8 (R) | 1024 | |
| Sputum | 1 Sp | ME, E, DO, SXT, AMC, DA, CIP | 24 | 16 |
| 2 Sp | ME, CIP | 17 | 128 | |
| Diabetic foot | 1 Df | ME, DO | 20 | 64 |
| Burn swab | 1 Bs | ME, E, DO, SXT, AMC, DA, CIP | 16 | 256 |
| Blood | 1 Bl | ME | 16 | 256 |
| Cerebrospinal fluid | 1 CSF | ME | 18 | 128 |
| Parameter | Experimental Group | p Value | SEM | |||
|---|---|---|---|---|---|---|
| Control | NSEI | NSEII | NSEIII | |||
| Initial body weight | 804 | 808 | 766 | 806 | 0.06 | 10.29 |
| Growing period (30–60 d) | ||||||
| Body weight, g | 2036 c | 2060 c | 2237 b | 2346 a | <0.03 | 15.14 |
| BWG, g | 1232 c | 1251 c | 1471 b | 1537 a | <0.04 | 18.16 |
| FI, g | 3033 b | 3030 b | 3372 a | 3351 a | 0.04 | 20.83 |
| FCR | 2.46 a | 2.42 a | 2.30 b | 2.18 c | 0.001 | <0.001 |
| Digestive enzymes | ||||||
| Chymotrypsin (U/gprot) | 69.96 c | 79.99 b | 83.69 ab | 85.36 a | 0.03 | 0.15 |
| Amylase (U/gprot) | 1.95 c | 2.10 c | 2.56 b | 2.96 a | 0.001 | 0.30 |
| Lipase (U/gprot) | 39.64 d | 42.55 c | 47.6 b | 49.67 a | 0.04 | 0.22 |
| Sample Source | MRSA Code No. | Serum Bactericidal Capacity (%) | Bactericidal Index | |
|---|---|---|---|---|
| Control | NSE Fed Rabbits | |||
| Sputum | 1 Sp | 43 | 86.1 | 43.2 |
| Pus | 1 Pu | 81.0 | 32.0 | |
| Sausage | 5 Sg | 74.2 | 31.1 | |
| Cow milk | 4 M | 55.0 | 1.2 | |
| Pus | 3 Pu | 50.0 | 0.7 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elmowalid, G.A.E.; Ahmad, A.A.M.; El-Hamid, M.I.A.; Ibrahim, D.; Wahdan, A.; El Oksh, A.S.A.; Yonis, A.E.; Elkady, M.A.; Ismail, T.A.; Alkhedaide, A.Q.; et al. Nigella sativa Extract Potentially Inhibited Methicillin Resistant Staphylococcus aureus Induced Infection in Rabbits: Potential Immunomodulatory and Growth Promoting Properties. Animals 2022, 12, 2635. https://doi.org/10.3390/ani12192635
Elmowalid GAE, Ahmad AAM, El-Hamid MIA, Ibrahim D, Wahdan A, El Oksh ASA, Yonis AE, Elkady MA, Ismail TA, Alkhedaide AQ, et al. Nigella sativa Extract Potentially Inhibited Methicillin Resistant Staphylococcus aureus Induced Infection in Rabbits: Potential Immunomodulatory and Growth Promoting Properties. Animals. 2022; 12(19):2635. https://doi.org/10.3390/ani12192635
Chicago/Turabian StyleElmowalid, Gamal Abd Elmoneim, Adel Attia M. Ahmad, Marwa I. Abd El-Hamid, Doaa Ibrahim, Ali Wahdan, Amal S. A. El Oksh, Ahlam E. Yonis, Mohamed Abdelrazek Elkady, Tamer Ahmed Ismail, Adel Qlayel Alkhedaide, and et al. 2022. "Nigella sativa Extract Potentially Inhibited Methicillin Resistant Staphylococcus aureus Induced Infection in Rabbits: Potential Immunomodulatory and Growth Promoting Properties" Animals 12, no. 19: 2635. https://doi.org/10.3390/ani12192635
APA StyleElmowalid, G. A. E., Ahmad, A. A. M., El-Hamid, M. I. A., Ibrahim, D., Wahdan, A., El Oksh, A. S. A., Yonis, A. E., Elkady, M. A., Ismail, T. A., Alkhedaide, A. Q., & Elnahriry, S. S. (2022). Nigella sativa Extract Potentially Inhibited Methicillin Resistant Staphylococcus aureus Induced Infection in Rabbits: Potential Immunomodulatory and Growth Promoting Properties. Animals, 12(19), 2635. https://doi.org/10.3390/ani12192635