

CRFB5a, a Subtype of Japanese Eel (Anguilla japonica) Type I IFN Receptor, Regulates Host Antiviral and Antimicrobial Functions through Activation of IRF3/IRF7 and LEAP2
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Tissue Collection and Immune Challenge
2.2. Cloning of the Full-Length cDNA of CRFB5a and Bioinformatics Analysis
2.3. Subcellular Localization Analysis
2.4. Expression Analysis of CRFB5a by RT-qPCR
2.5. Luciferase Activity Assay
2.6. Western Blotting
2.7. Overexpression of CRFB5a
2.8. RNA Interference of CRFB5a In Vivo and In Vitro
2.9. Statistical Analysis
3. Results
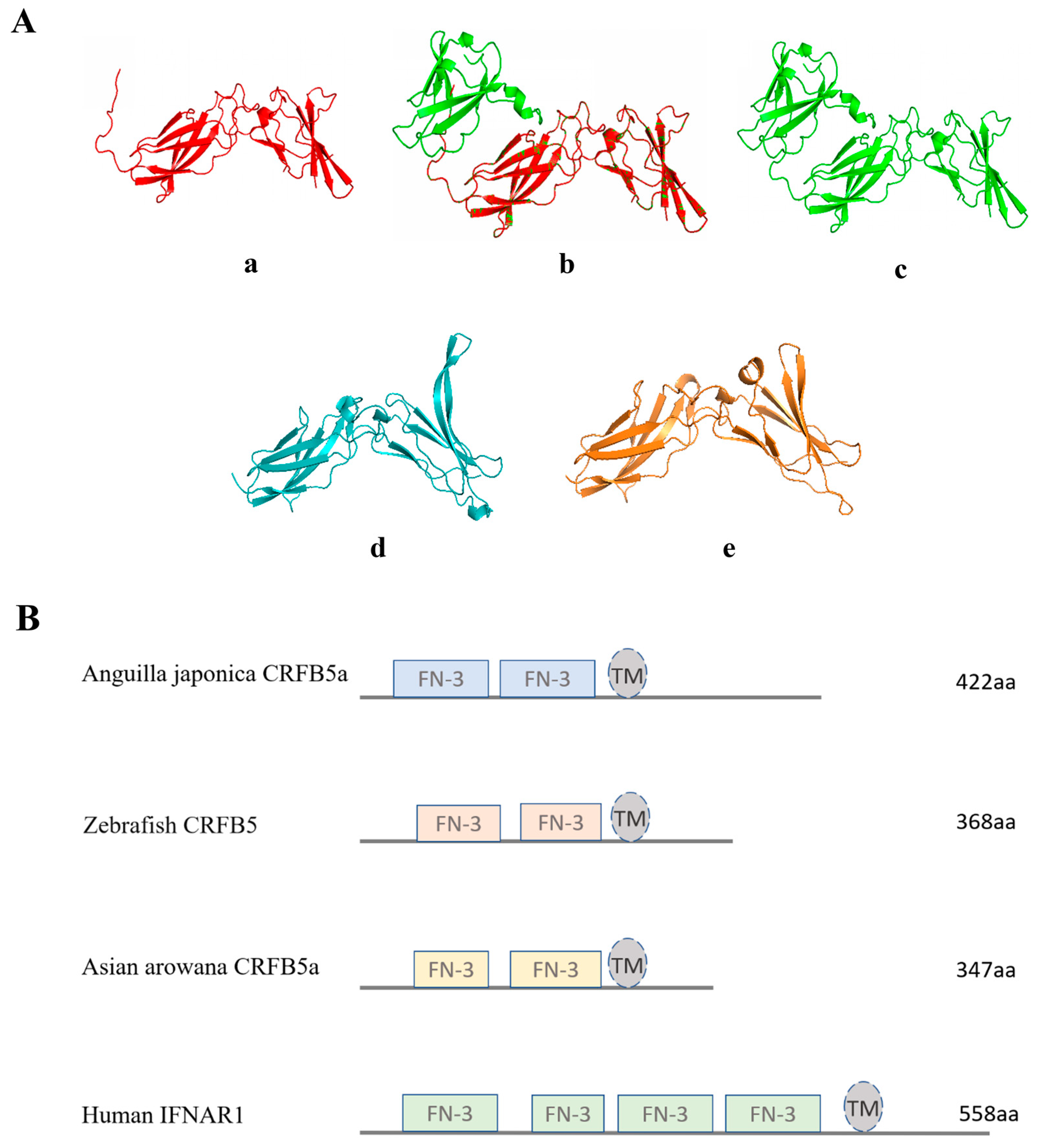
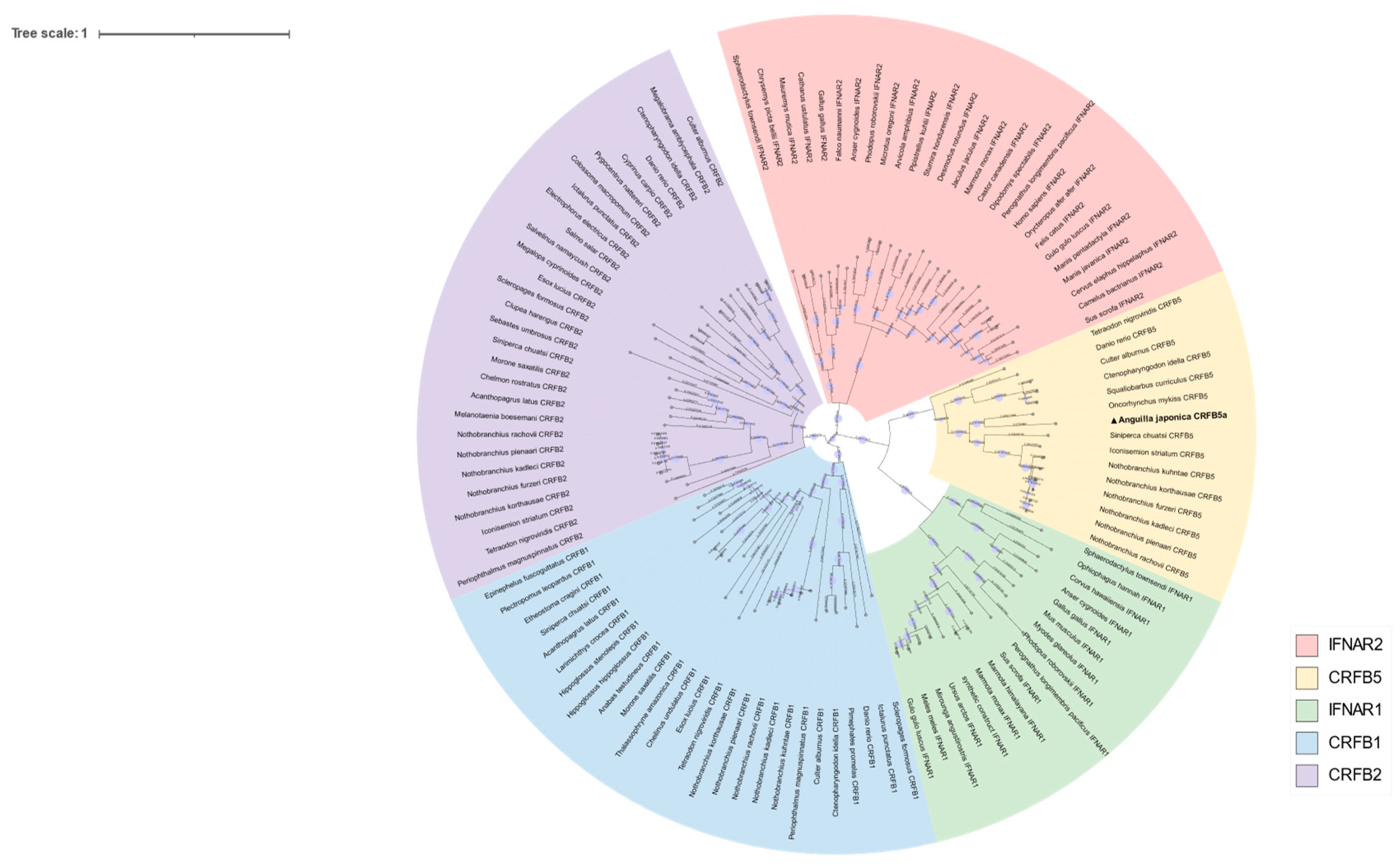
3.1. Sequence Analysis of CRFB5a from Japanese Eel
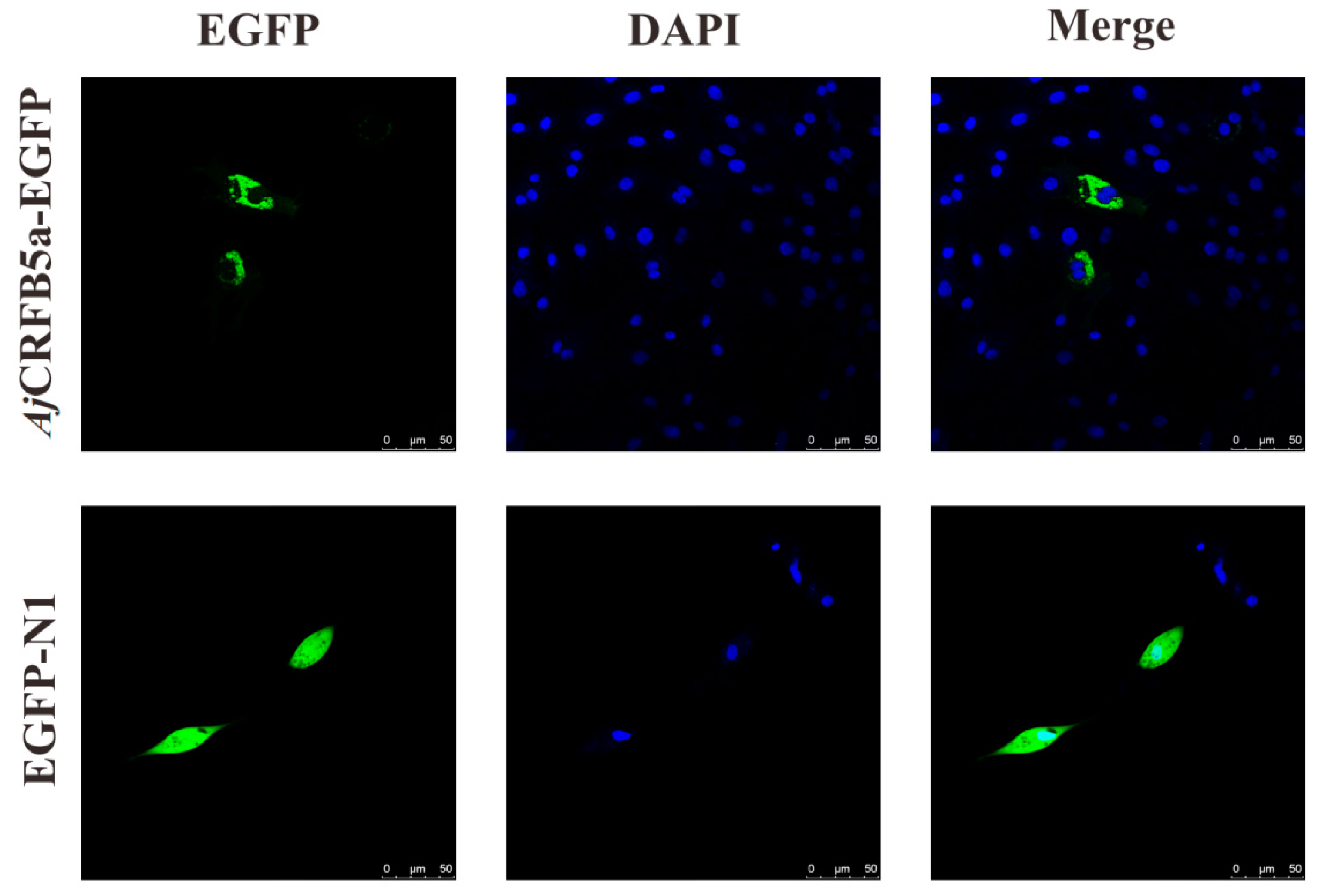
3.2. Subcellular Localization of AjCRFB5a
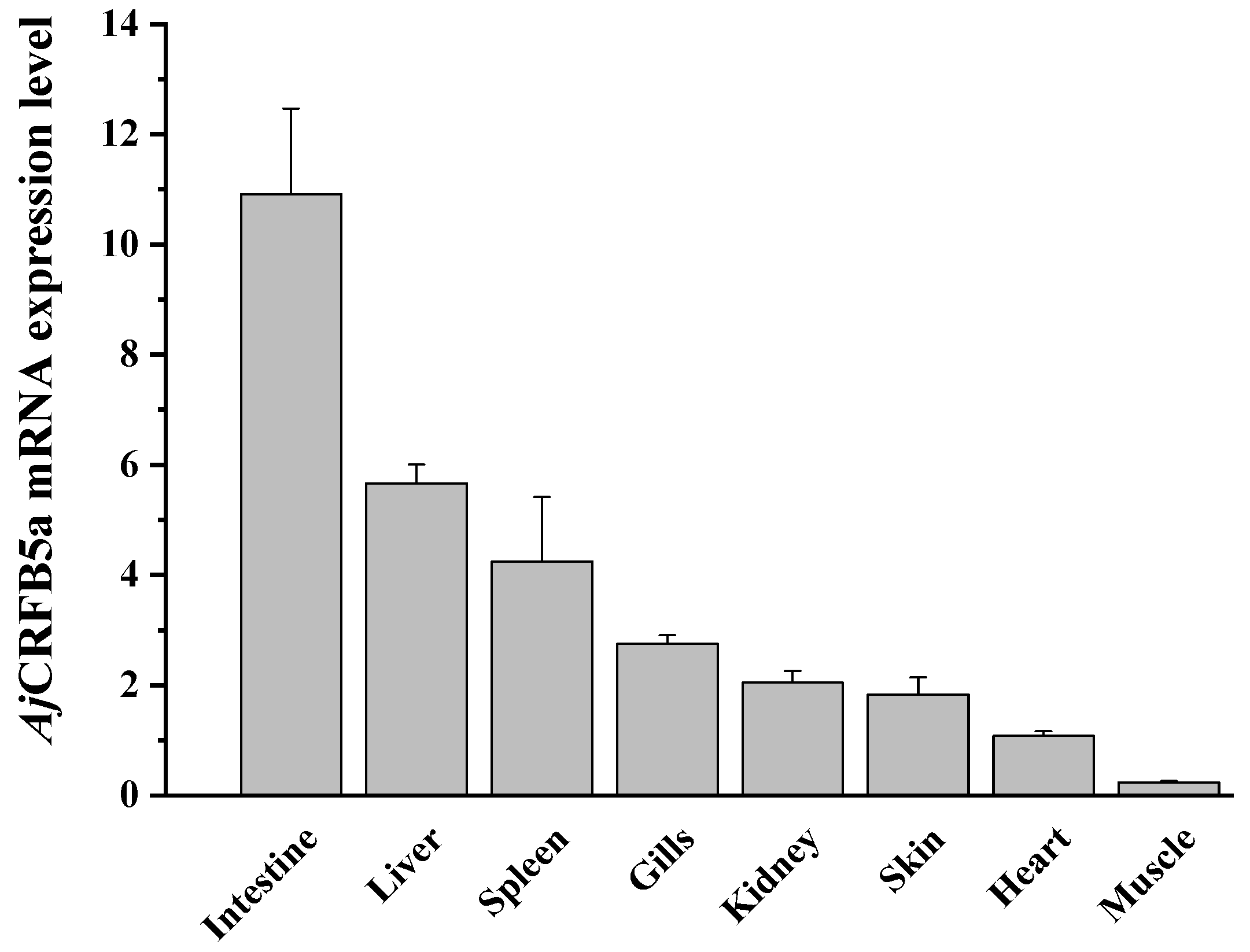
3.3. Tissue Expression of AjCRFB5a
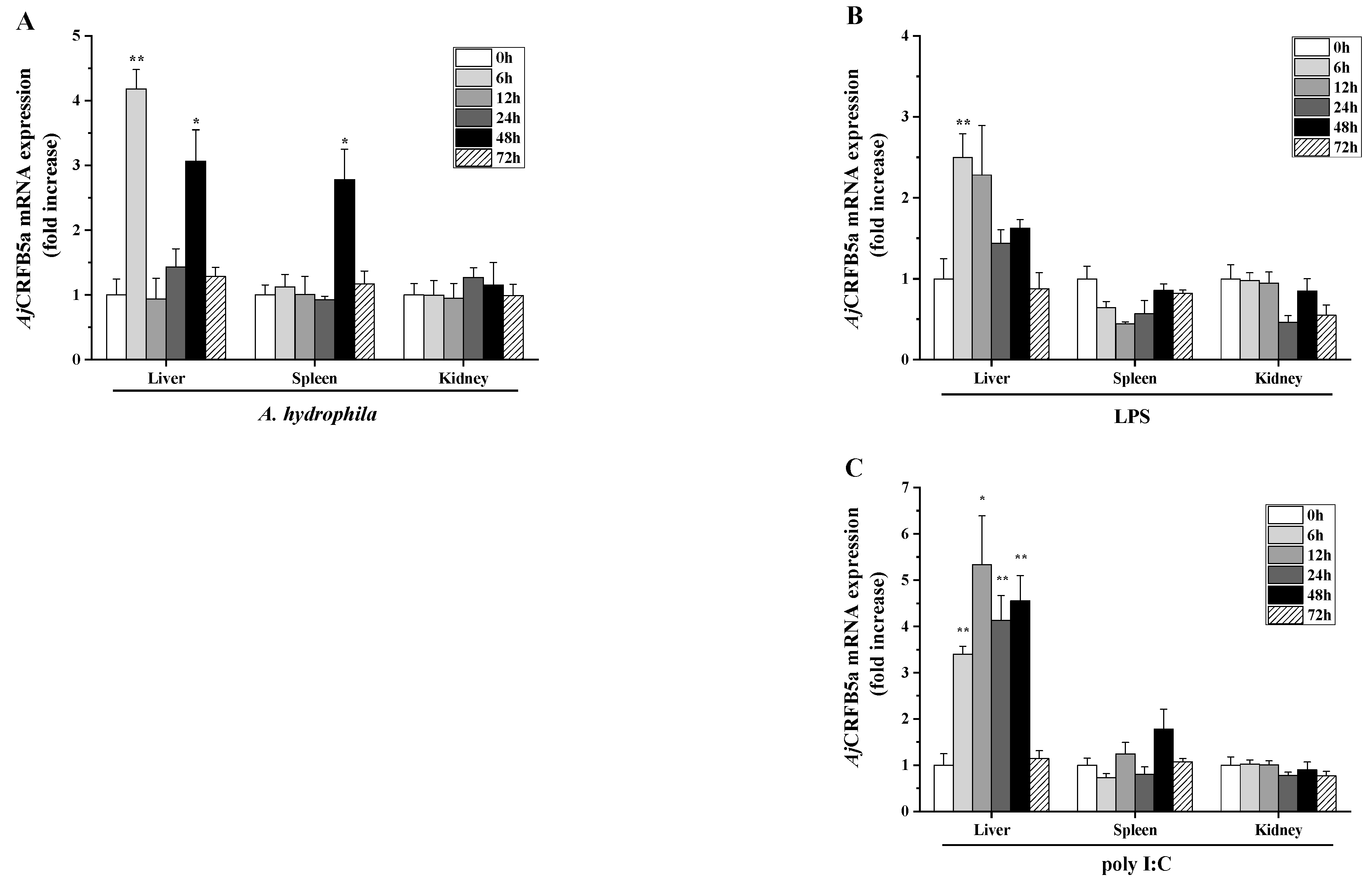
3.4. Gene Expression of AjCRFB5a In Vivo upon Stimulation
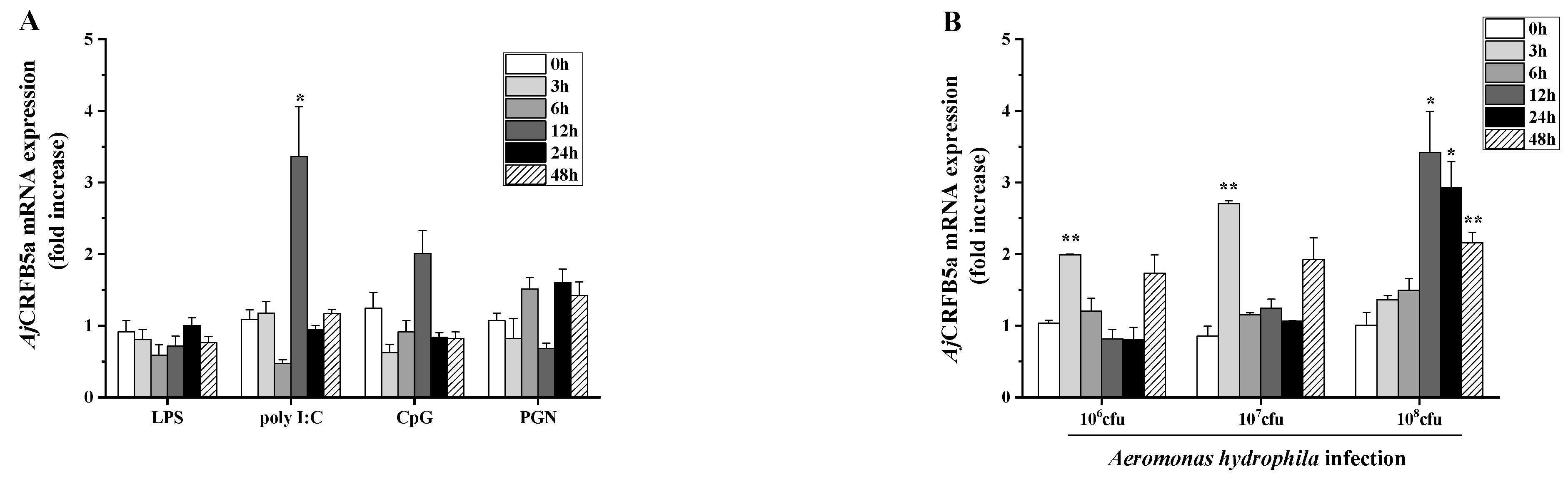
3.5. Gene Expression of AjCRFB5a In Vitro after Stimulation
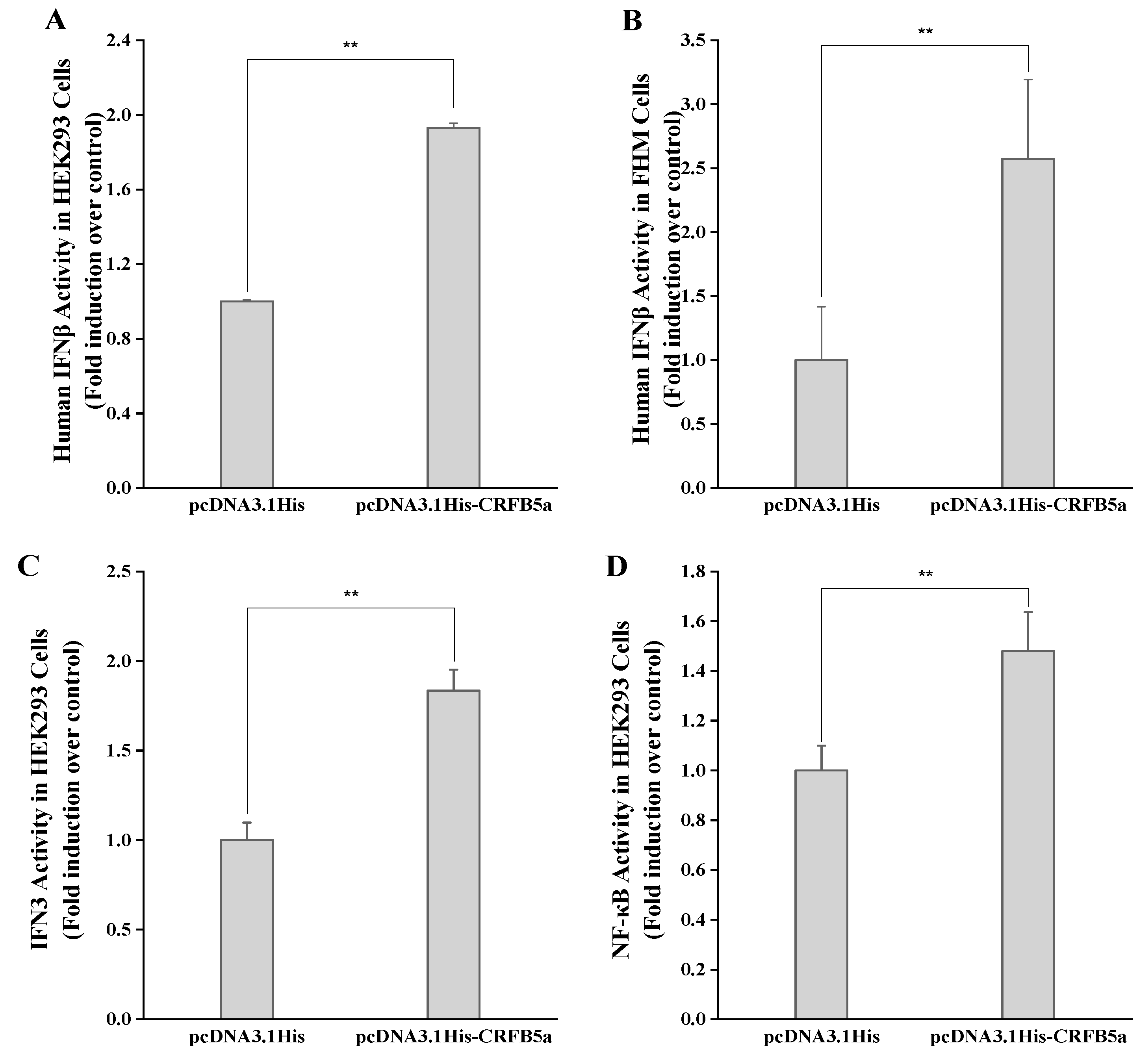
3.6. Dual Luciferase Activity of the AjCRFB5a Gene
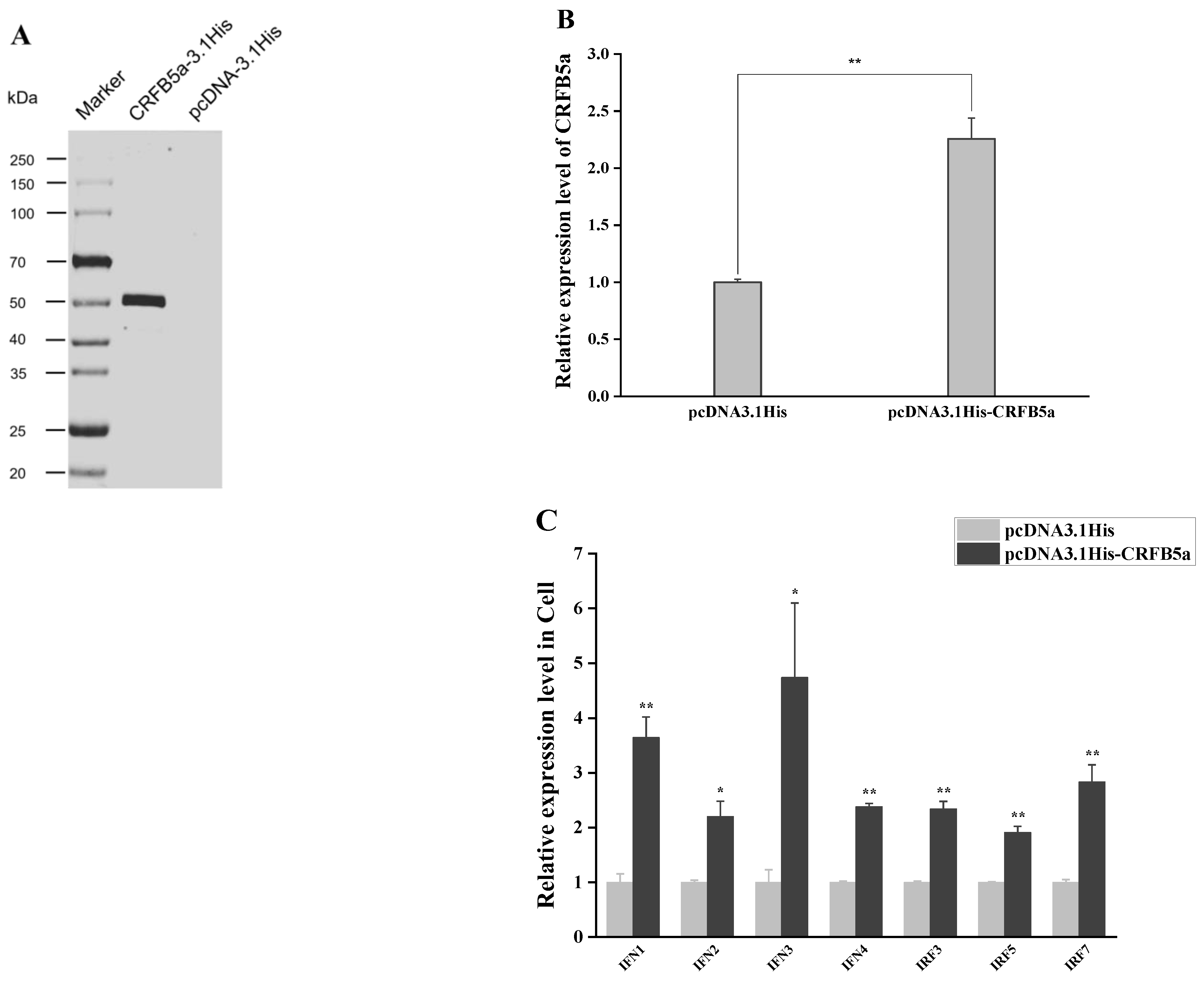
3.7. Induction of Immune-Related Genes under Overexpression of AjCRFB5a
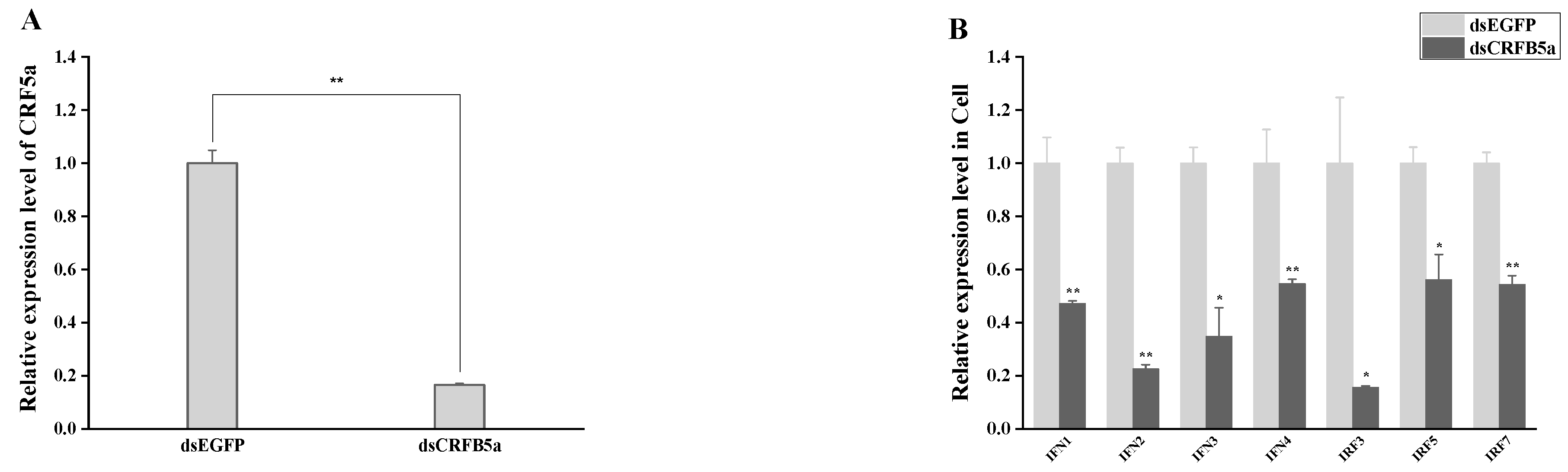
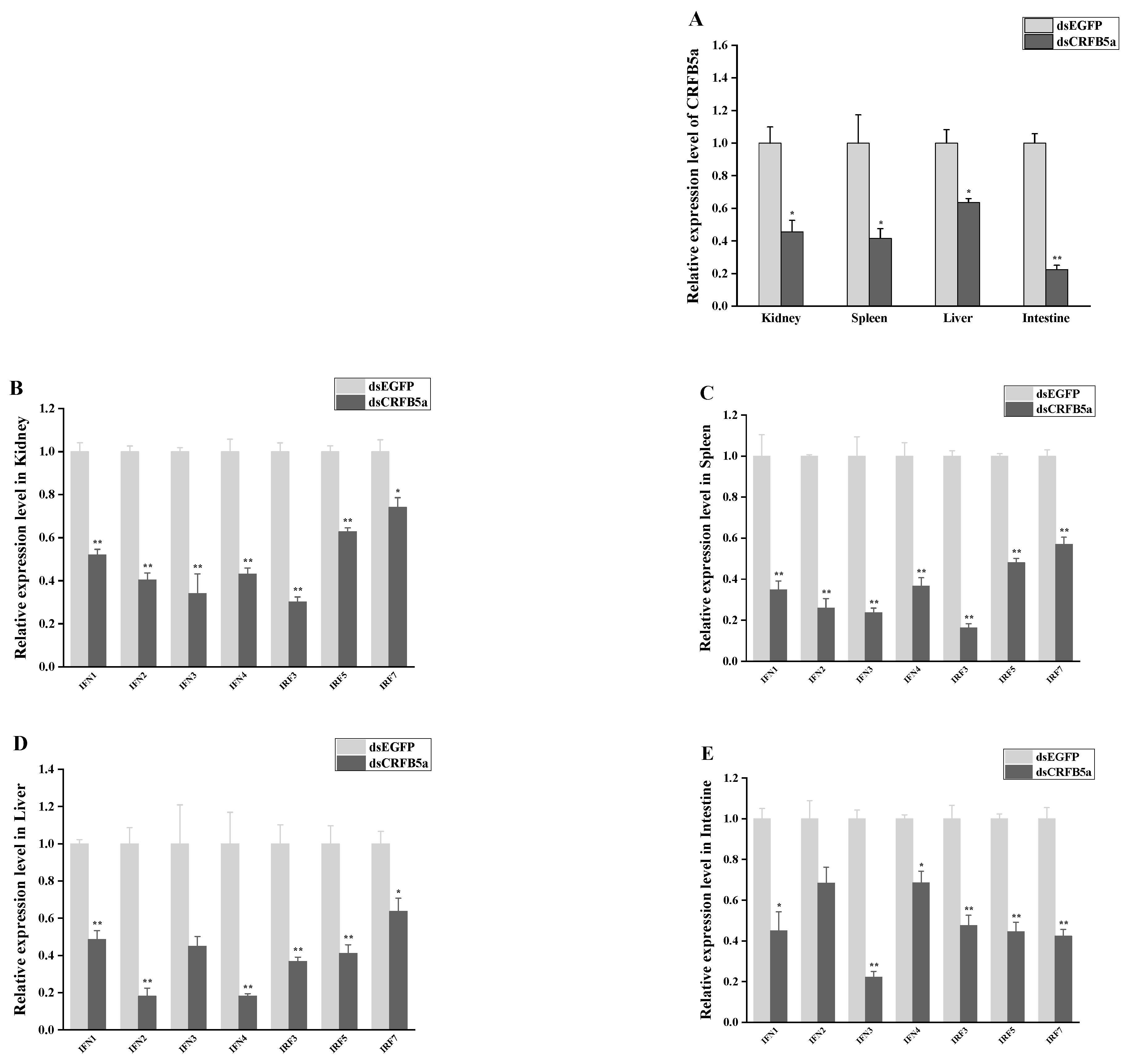
3.8. The effect of AjCRFB5a Gene Silencing on the Immune-Related Genes from Type I IFN Signaling Pathways
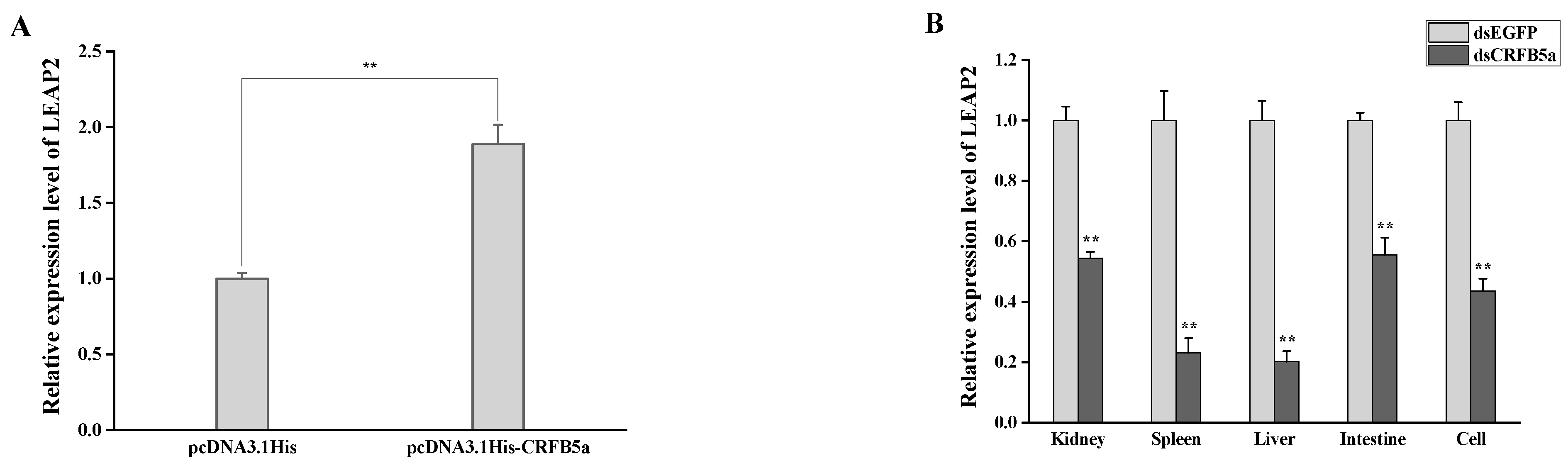
3.9. The Effect of AjCRFB5a on the Antimicrobial Peptide LEAP2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, S.; Lu, L.-F.; Feng, H.; Wu, N.; Chen, D.-D.; Zhang, Y.-B.; Gui, J.-F.; Nie, P.; Zhang, Y.-A. IFN Regulatory Factor 10 Is a Negative Regulator of the IFN Responses in Fish. J. Immunol. 2014, 193, 1100–1109. [Google Scholar] [CrossRef] [PubMed]
- Pellefigues, C.; Tang, S.-C.; Schmidt, A.; White, R.F.; Lamiable, O.; Connor, L.M.; Ruedl, C.; Dobrucki, J.; Le Gros, G.; Ronchese, F. Toll-Like Receptor 4, but Not Neutrophil Extracellular Traps, Promote IFN Type I Expression to Enhance Th2 Responses to Nippostrongylus brasiliensis. Front. Immunol. 2017, 8, 1575. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Bols, N.C.; Pham, P.H.; Secombes, C.J.; Zou, J. Evolution of IFN subgroups in bony fish—1: Group I-III IFN exist in early ray-finned fish, with group II IFN subgroups present in the Holostean spotted gar, Lepisosteus oculatus. Fish Shellfish Immunol. 2019, 95, 163–170. [Google Scholar] [CrossRef]
- Li, L.; Chen, S.N.; Laghari, Z.A.; Huo, H.J.; Hou, J.; Huang, L.; Li, N.; Nie, P. Myxovirus resistance (Mx) gene and its differential expression regulated by three type I and two type II IFNs in mandarin fish, Siniperca chuatsi. Dev. Comp. Immunol. 2020, 105, 103604. [Google Scholar] [CrossRef]
- Wei, L.; Zhang, R.; Zhang, J.; Li, J.; Kong, D.; Wang, Q.; Fang, J.; Wang, L. PRKAR2A deficiency protects mice from experimental colitis by increasing IFN-stimulated gene expression and modulating the intestinal microbiota. Mucosal Immunol. 2021, 14, 1282–1294. [Google Scholar] [CrossRef] [PubMed]
- Duncan, C.J.; Randall, R.E.; Hambleton, S. Genetic Lesions of Type I Interferon Signalling in Human Antiviral Immunity. Trends Genet. 2021, 37, 46–58. [Google Scholar] [CrossRef]
- Gui, J.; Zhao, B.; Lyu, K.; Tong, W.; Fuchs, S.Y. Downregulation of the IFNAR1 chain of type 1 interferon receptor contributes to the maintenance of the haematopoietic stem cells. Cancer Biol. Ther. 2017, 18, 534–543. [Google Scholar] [CrossRef]
- Walter, M.R. The Role of Structure in the Biology of Interferon Signaling. Front. Immunol. 2020, 11, 606489. [Google Scholar] [CrossRef]
- Langevin, C.; Boudinot, P.; Collet, B. IFN Signaling in Inflammation and Viral Infections: New Insights from Fish Models. Viruses 2019, 11, 302. [Google Scholar] [CrossRef]
- Liu, Y.; Du, H.; Wang, S.; Lv, Y.; Deng, H.; Chang, K.; Zhou, P.; Hu, C. Grass carp (Ctenopharyngodon idella) TNK1 modulates JAK-STAT signaling through phosphorylating STAT1. Dev. Comp. Immunol. 2021, 116, 103951. [Google Scholar] [CrossRef]
- Gong, X.-Y.; Zhang, Q.-M.; Gui, J.-F.; Zhang, Y.-B. SVCV infection triggers fish IFN response through RLR signaling pathway. Fish Shellfish Immunol. 2019, 86, 1058–1063. [Google Scholar] [CrossRef] [PubMed]
- Lutfalla, G.; Crollius, H.R.; Stange-Thomann, N.; Jaillon, O.; Mogensen, K.; Monneron, D. Comparative genomic analysis reveals independent expansion of a lineage-specific gene family in vertebrates: The class II cytokine receptors and their ligands in mammals and fish. BMC Genom. 2003, 4, 29. [Google Scholar] [CrossRef]
- Chen, Y.-M.; Kuo, C.-E.; Chen, G.-R.; Kao, Y.-T.; Zou, J.; Secombes, C.J.; Chen, T.-Y. Functional analysis of an orange-spotted grouper (Epinephelus coioides) interferon gene and characterisation of its expression in response to nodavirus infection. Dev. Comp. Immunol. 2014, 46, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Gan, Z.; Cheng, J.; Chen, S.; Hou, J.; Li, N.; Xia, H.; Xia, L.; Lu, Y.; Nie, P. Identification and characterization of tilapia CRFB1, CRFB2 and CRFB5 reveals preferential receptor usage of three IFN subtypes in perciform fishes. Fish Shellfish Immunol. 2020, 107, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, W.; Wang, B.; Mao, H.; Sun, Z.; Hou, Q.; Mi, Y.; Fan, L.; Hu, C. Cloning, identification of the two cytokine receptor family B subunits CRFB1 and CRFB5 from grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2015, 45, 211–220. [Google Scholar] [CrossRef]
- Sun, B.; Greiner-Tollersrud, L.; Koop, B.F.; Robertsen, B. Atlantic salmon possesses two clusters of type I interferon receptor genes on different chromosomes, which allows for a larger repertoire of interferon receptors than in zebrafish and mammals. Dev. Comp. Immunol. 2014, 47, 275–286. [Google Scholar] [CrossRef]
- Pang, A.N.; Chen, S.N.; Gan, Z.; Li, L.; Li, N.; Wang, S.; Sun, Z.; Liu, L.H.; Sun, Y.L.; Song, X.J.; et al. Identification of type II interferons and receptors in an osteoglossiform fish, the arapaima Arapaima gigas. Dev. Comp. Immunol. 2023, 139, 104589. [Google Scholar] [CrossRef]
- Tang, Z.Z.; Wang, T.-Y.; Chen, Y.-M.; Chen, T.-Y. Cloning and characterisation of type I interferon receptor 1 in orange-spotted grouper (Epinephelus coioides) for response to nodavirus infection. Fish Shellfish Immunol. 2020, 101, 302–311. [Google Scholar] [CrossRef]
- Stein, C.; Caccamo, M.; Laird, G.; Leptin, M. Conservation and divergence of gene families encoding components of innate immune response systems in zebrafish. Genome Biol. 2007, 8, R251. [Google Scholar] [CrossRef]
- Wang, S.; Chen, S.N.; Sun, Z.; Pang, A.N.; Wang, S.; Liu, L.H.; Liu, Y.; Nie, P. Four type I IFNs, IFNa1, IFNa2, IFNb, IFNc, and their receptor usage in an osteoglossomorph fish, the Asian arowana, Scleropages formosus. Fish Shellfish Immunol. 2021, 117, 70–81. [Google Scholar] [CrossRef]
- Liao, Z.; Wan, Q.; Su, J. Bioinformatics analysis of organizational and expressional characterizations of the IFNs, IRFs and CRFBs in grass carp Ctenopharyngodon idella. Dev. Comp. Immunol. 2016, 61, 97–106. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, J.; Feng, J.; Wu, K.; Chen, K.; Jia, Z.; Zhu, X.; Huang, W.; Zhao, X.; Liu, Q.; et al. Structural and Functional Analyses of Type I IFNa Shed Light into Its Interaction with Multiple Receptors in Fish. Front. Immunol. 2022, 13, 862764. [Google Scholar] [CrossRef]
- Takeuchi, A.; Watanabe, S.; Yamamoto, S.; Miller, M.; Fukuba, T.; Miwa, T.; Okino, T.; Minamoto, T.; Tsukamoto, K. First use of oceanic environmental DNA to study the spawning ecology of the Japanese eel Anguilla japonica. Mar. Ecol. Prog. Ser. 2019, 609, 187–196. [Google Scholar] [CrossRef]
- Itakura, H.; Miyake, Y.; Kitagawa, T.; Kimura, S. Site fidelity, diel and seasonal activities of yellow-phase Japanese eels (Anguilla japonica) in a freshwater habitat as inferred from acoustic telemetry. Ecol. Freshw. Fish 2018, 27, 737–751. [Google Scholar] [CrossRef]
- Mizutani, T.; Sayama, Y.; Nakanishi, A.; Ochiai, H.; Sakai, K.; Wakabayashi, K.; Tanaka, N.; Miura, E.; Oba, M.; Kurane, I.; et al. Novel DNA virus isolated from samples showing endothelial cell necrosis in the Japanese eel, Anguilla japonica. Virology 2011, 412, 179–187. [Google Scholar] [CrossRef]
- Schulz, P.; Kaczorek, E.; Małaczewska, J.; Siwicki, A. Immunopathogenesis of Herpesviruses: Influence of Anguillid Herpes Virus 1 on Macrophage and Lymphocyte Activity in European eel (Anguilla anguilla). J. Comp. Pathol. 2016, 154, 97. [Google Scholar] [CrossRef]
- Feng, J.J.; Xu, Y.K.; Lin, P.; Wang, Y.L.; Zhang, Z.P.; Zou, P.F.; Peng, X.W. Fish IKK alpha from Japanese eel (Anguilla japonica) can activate NF-kappa B, AP1, and type I IFN signaling pathways. Fish Shellfish Immunol. 2020, 106, 982–992. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Zhong, J.; Zhang, Z.; Zou, P.; Wang, Y. Comparative Transcriptome Reveals the Potential Modulation Mechanisms of Spfoxl-2 Affecting Ovarian Development of Scylla paramamosain. Mar. Biotechnol. 2022, 24, 125–135. [Google Scholar] [CrossRef]
- Luo, K.; Zhang, S.; Tang, D.; Xia, L.; Gao, W.; Tian, G.; Qi, Z.; Xu, Q.; Zhang, W. Analysis of the expression patterns of the cytokine receptor family B (CRFB) and interferon gamma receptor (IFNGR) in Dabry’s sturgeon (Acipenser dabryanus). Dev. Comp. Immunol. 2018, 84, 420–426. [Google Scholar]
- Kato, H.; Takeuchi, O.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Matsui, K.; Uematsu, S.; Jung, A.; Kawai, T.; Ishii, K.J.; et al. Differential roles of MDA5 and RIG-I hel-icases in the recognition of RNA viruses. Nature 2006, 441, 101–105. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, S.; Mao, Y.; Qiao, G.; Li, Q. Identification and analysis of differentially expressed microRNAs in gibel carp Carassius auratus gibelio responding to polyinosinic-polycytidylic acid (poly I:C) stimulation. Fish Shellfish Immunol. Rep. 2023, 4, 100083. [Google Scholar] [CrossRef]
- Li, L.; Chen, S.N.; Laghari, Z.A.; Huang, B.; Huo, H.J.; Li, N.; Nie, P. Receptor complex and signalling pathway of the two type II IFNs, IFN-gamma and IFN-gamma rel in mandarin fish or the so-called Chinese perch Siniperca chuatsi. Dev. Comp. Immunol. 2019, 97, 98–112. [Google Scholar] [CrossRef] [PubMed]
- Rathinam, V.A.; A Fitzgerald, K. Cytosolic surveillance and antiviral immunity. Curr. Opin. Virol. 2011, 1, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Wang, Z.; Liang, Y.; Zhai, S.; Huang, W.; Nie, P. Identification of four type I IFNs from Japanese eel with differential expression properties and Mx promoter inducibility. Dev. Comp. Immunol. 2019, 91, 62–71. [Google Scholar] [CrossRef]
- Zhang, Q.; Lenardo, M.J. Baltimore, 30 Years of NF-kappaB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-H.; Sharrocks, A.D.; Whitmarsh, A.J. MAP kinase signalling cascades and transcriptional regulation. Gene 2013, 513, 1–13. [Google Scholar] [CrossRef]
- Duan, M.Z.; Huang, B.; Liang, Y.; Zhang, F.F.; Nie, P.; Huang, W.S. Molecular cloning and expression analysis of a liver expressed antimicrobial peptide-2 in Japanese eel, Anguilla japonica. Acta Hydrobiol. Sin. 2016, 40, 252–260. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence |
|---|---|
| Specific primers for head-to-toe PCR | |
| CRFB5a-ORF5′ | 5′-ATGAATGTAGGCCTTGGGGTTG-3′ |
| CRFB5a-ORF3′ | 5′-GACACACTCCTCCAGAATGTCCTC-3′ |
| Specific primers for 5′ RACE | |
| CRFB5a-5-inner | 5′-GTGAAGGTGATGCTCTGATTGGC-3′ |
| CRFB5a-5-outer | 5′-CGGAGGGGAAGAAGGTGGATTTGA-3′ |
| Specific primers for RT-qPCR | |
| 5′real-CRFB5a | 5′-CGAGGTTTGAACACGAGCATC-3′ |
| 3′real-CRFB5a | 5′-TCTGCTGGCACTGCGTTTC-3′ |
| 5′real-IFN1 | 5′-CTTGCAGGTTGAGGAACGCAT-3′ |
| 3′real-IFN1 | 5′-GCATTCTTTCAGCTCCGAAGC-3′ |
| 5′real-IFN2 | 5′-CTCAGTGAGATGGGTGGAGA-3′ |
| 3′real-IFN2 | 5′-GCTCGCAGAAGAAACACATTTC-3′ |
| 5′real-IFN3 | 5′-TGACATCACGAGCAAATCTCAG-3′ |
| 3′real-IFN3 | 5′-CTCGAACACACTGCTCCAAGT-3′ |
| 5′real-IFN4 | 5′-GCTGGGCAGAGCGACATCATT-3′ |
| 3′real-IFN4 | 5′-AGAATCGTCACTCCCTGGCTT-3′ |
| 5′real-IRF3 | 5′-CCAGGCACACCAAGCGAGTTC-3′ |
| 3′real-IRF3 | 5′-ACCTCTTGAGGAATGTTGCTGTTCG-3′ |
| 5′real-IRF5 | 5′-CCAGGTGATTCCCGTCGTTGC-3′ |
| 3′real-IRF5 | 5′-CCAGGTGATTCCCGTCGTTGC-3′ |
| 5′real-IRF7 | 5′-AGATGCGTATGCGACCGATTGC-3′ |
| 3′real-IRF7 | 5′-ATCATCCTGCTGCTGGTTGTTCAG-3′ |
| 5′real-LEAP-2 | 5′-ACTGCCTGCGGTTTGGTGTTG-3′ |
| 3′real-LEAP-2 | 5′-GCTTGCTTCCCATGATCCTCCAC-3′ |
| Specific primers for dsRNA preparation | |
| 5′dsCRFB5a | 5′-TGGGGTTGCTGTTATTGTCA-3′ |
| 3′dsCRFB5a | 5′-TGGGGTTGCTGTTATTGTCA-3′ |
| 5′dsCRFB5a-T7 | 5′-TAATACGACTCACTATAGGGTGGGGTTGCTGTTATTGTCA-3′ |
| 3′dsCRFB5a-T7 | 5′-TAATACGACTCACTATAGGGTGGGGTTGCTGTTATTGTCA-3′ |
| 5′dsEGFP | 5′-GGTGAACTTCAAGATCCGCC-3′ |
| 3′dsEGFP | 5′-CTTGTACAGCTCGTCCATGC-3′ |
| 5′dsEGFP-T7 | 5′-TAATACGACTCACTATAGGGGGTGAACTTCAAGATCCGCC-3′ |
| 3′dsEGFP-T7 | 5′-TAATACGACTCACTATAGGGCTTGTACAGCTCGTCCATGC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Lin, P.; Wang, Y.; Lai, X.; Chen, P.; Li, F.; Feng, J. CRFB5a, a Subtype of Japanese Eel (Anguilla japonica) Type I IFN Receptor, Regulates Host Antiviral and Antimicrobial Functions through Activation of IRF3/IRF7 and LEAP2. Animals 2023, 13, 3157. https://doi.org/10.3390/ani13193157
Wang T, Lin P, Wang Y, Lai X, Chen P, Li F, Feng J. CRFB5a, a Subtype of Japanese Eel (Anguilla japonica) Type I IFN Receptor, Regulates Host Antiviral and Antimicrobial Functions through Activation of IRF3/IRF7 and LEAP2. Animals. 2023; 13(19):3157. https://doi.org/10.3390/ani13193157
Chicago/Turabian StyleWang, Tianyu, Peng Lin, Yilei Wang, Xiaojian Lai, Pengyun Chen, Fuyan Li, and Jianjun Feng. 2023. "CRFB5a, a Subtype of Japanese Eel (Anguilla japonica) Type I IFN Receptor, Regulates Host Antiviral and Antimicrobial Functions through Activation of IRF3/IRF7 and LEAP2" Animals 13, no. 19: 3157. https://doi.org/10.3390/ani13193157
APA StyleWang, T., Lin, P., Wang, Y., Lai, X., Chen, P., Li, F., & Feng, J. (2023). CRFB5a, a Subtype of Japanese Eel (Anguilla japonica) Type I IFN Receptor, Regulates Host Antiviral and Antimicrobial Functions through Activation of IRF3/IRF7 and LEAP2. Animals, 13(19), 3157. https://doi.org/10.3390/ani13193157