
Identification of Genetic Polymorphisms of PI, PIII, and Exon 53 in the Acetyl-CoA Carboxylase-α (ACACα) Gene and Their Association with Milk Composition Traits of Najdi Sheep

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Management, and Experimental Design
2.2. Collection and Analysis of Milk Samples
2.3. Blood Sampling and DNA Extraction
2.4. Amplification of the ACACα Gene via Polymerase Chain Reaction (PCR)
2.5. Sequencing and Genotype Analysis
2.6. Analyses of the Associations and Effects of Alleles
2.7. Linkage Disequilibrium (LD) Estimation
3. Results
3.1. Sequence Analysis of the ACACα Gene Coding Region
3.2. Association of Genotype of ACACα Gene with Milk Traits
4. Discussion
4.1. Sequence Analysis of the ACACα Gene Coding Region
4.2. Association of Genotype of ACACα Gene with Milk Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Travers, M.T.; Vallance, A.J.; Gourlay, H.T.; Gill, C.A.; Klein, I.; Bottema, C.B.K.; Barber, M.C. Promoter I of the ovine acetyl-CoA carboxylase-α gene: An E-box motif at −114 in the proximal promoter binds upstream stimulatory factor (USF)-1 and USF-2 and acts as an insulin-response sequence in differentiating adipocytes. Biochem. J. 2001, 359, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Micha, R.; Wallace, S. Effects on coronary heart disease of increasing polyunsaturated fat in place of saturated fat: A systematic review and meta-analysis of randomized controlled trials. PLoS Med 2010, 7, e1000252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kęsek, M.; Szulc, T.; Zielak-Steciwko, A. Reports Genetic, physiological and nutritive factors affecting the fatty acid profile in cows’ milk–a review. Anim. Sci. Pap. 2014, 32, 95–105. [Google Scholar]
- Ramsay, R.R.; Gandour, R.D.; van der Leij, F.R. Molecular enzymology of carnitine transfer and transport. Biochim. Biophys. Acta 2001, 1546, 21–43. [Google Scholar] [CrossRef]
- Brownsey, R.W.; Boone, A.N.; Elliott, J.E.; Kulpa, J.E.; Lee, W.M. Regulation of acetyl-CoA carboxylase. Biochem. Soc. Trans. 2006, 34, 223–227. [Google Scholar] [CrossRef]
- Travers, M.T.; Barber, M.C. Acetyl-CoA carboxylase-α: Gene structure-function relationships. J. Anim. Sci. 2001, 79, E136–E143. [Google Scholar] [CrossRef]
- García-Fernández, M.; Sánchez, J.; Gutiérrez-Gil, B.; García-Gámez, E.; Arranz, J. Association between acetyl-CoA carboxylase α (ACACA) SNPs and milk fatty acid profile in Spanish Churra sheep. In Proceedings of the 9th World Congress on Genetics Applied to Livestock Production, Dept. Produccion Animal, Universidad de Leon, Leon, Spain, 1–6 August 2010. [Google Scholar]
- Lopez-Casillas, F.; Ponce-Castaneda, M.V.; Kim, K.H. In vivo regulation of the activity of the two promoters of the rat acetyl coenzyme-A carboxylase gene. Endocrinology 1991, 129, 1049–1058. [Google Scholar] [CrossRef]
- Barber, M.C.; Travers, M.T. Elucidation of a promoter activity that directs the expression of acetyl-CoA carboxylase alpha with an alternative N-terminus in a tissue-restricted fashion. Biochem. J. 1998, 333 Pt 1, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Federica, S.; Francesco, N.; Giovanna de, M.; Carmela, S.M.; Gennaro, C.; Carmela, T.; Bianca, M. Identification of novel single nucleotide polymorphisms in promoter III of the acetyl-CoA carboxylase-alpha gene in goats affecting milk production traits. J. Hered. 2009, 100, 386–389. [Google Scholar] [CrossRef]
- Gerlando, R.D.; Mastrangelo, S.; Tortorici, L.; Tolone, M.; Sutera, A.M.; Sardina, M.T.; Portolano, B. Full-length sequencing and identification of novel polymorphisms in the ACACA gene of Valle del Belice sheep breed. J. Genet. 2017, 96, 591–597. [Google Scholar] [CrossRef] [Green Version]
- Mao, J.; Marcos, S.; Davis, S.K.; Burzlaff, J.; Seyfert, H.-M. Genomic distribution of three promoters of the bovine gene encoding acetyl-CoA carboxylase α and evidence that the nutritionally regulated promoter I contains a repressive element different from that in rat. Biochem. J. 2001, 358, 127–135. [Google Scholar] [CrossRef]
- Ayadi, M.; Matar, A.; Aljumaah, R.; Alshaikh, M.; Abouheif, M. Factors affecting milk yield, composition and udder health of Najdi ewes. Int. J. Anim. Vet. Adv. 2014, 6, 28–33. [Google Scholar] [CrossRef]
- Luna, P.; Juarez, M.; de la Fuente, M.A. Validation of a rapid milk fat separation method to determine the fatty acid profile by gas chromatography. J. Dairy Sci. 2005, 88, 3377–3381. [Google Scholar] [CrossRef] [Green Version]
- Sbihi, H.M.; Nehdi, I.A.; Tan, C.P.; Al-Resayes, S.I. Characteristics and fatty acid composition of milk fat from Saudi Aradi goat. Grasas Y Aceites 2015, 66, 101. [Google Scholar] [CrossRef] [Green Version]
- Matar, A.M.; Ayadi, M.; Aljumaah, R.S.; Nehdi, I.A.; Sbihi, H.M.; Souli, A.; Abouheif, M.A. Changes in the composition and fatty acid profile of Najdi ewes’ milk before and after weaning. S. Afr. J. Anim. Sci. 2017, 47, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Moioli, B.; Napolitano, F.; Orrù, L.; Catillo, G. Single nucleotide polymorphism detection in promoter I of the acetyl-CoA carboxylase-α gene in sheep. Small Rumin. Res. 2005, 59, 49–53. [Google Scholar] [CrossRef]
- Sztankoova, Z.; Rychtarova, J.; Kyselová, J.; Czerneková, V. Simultaneous genotyping of 4 SNPs in promoter III of the ovine ACACA. Small Rumin. Res. 2016, 138, 25–30. [Google Scholar] [CrossRef]
- Sherman, E.L.; Nkrumah, J.D.; Murdoch, B.M.; Li, C.; Wang, Z.; Fu, A.; Moore, S.S. Polymorphisms and haplotypes in the bovine neuropeptide Y, growth hormone receptor, ghrelin, insulin-like growth factor 2, and uncoupling proteins 2 and 3 genes and their associations with measures of growth, performance, feed efficiency, and carcass merit in beef cattle. J. Anim. Sci. 2008, 86, 1–16. [Google Scholar] [CrossRef]
- Hong, E.P.; Park, J.W. Sample size and statistical power calculation in genetic association studies. Genom. Inf. 2012, 10, 117–122. [Google Scholar] [CrossRef]
- Gauderman, W.J. Sample size requirements for association studies of gene-gene interaction. Am. J. Epidemiol. 2002, 155, 478–484. [Google Scholar] [CrossRef] [Green Version]
- Moioli, B.; Napolitano, F.; Orru, L.; Catillo, G. Single nucleotide polymorphism detection in promoter III of the acetyl-CoA carboxylase-alpha gene in sheep. J. Anim. Breed. Genet. 2005, 122, 418–420. [Google Scholar] [CrossRef] [PubMed]
- Moioli, B.; Scata, M.C.; De Matteis, G.; Annicchiarico, G.; Catillo, G.; Napolitano, F. The ACACA gene is a potential candidate gene for fat content in sheep milk. Anim. Genet. 2013, 44, 601–603. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, M.; Gutiérrez-Gil, B.; Garcia-Gámez, E.; Arranz, J.J. Identification of single nucleotide polymorphisms in the ovine acetyl-CoA carboxylase-alpha gene. Small Rumin. Res. 2010, 90, 34–40. [Google Scholar] [CrossRef]
- Singh, K.V.; Jayakumar, S.; Dixit, S.; Malik, Z. SNPs association studies in promoter III of the acetyl coenzyme-A carboxylase-α gene (ACACA) in Munjal–A threatened sheep population of India. Indian J. Anim. Sci. 2018, 88, 7. [Google Scholar] [CrossRef]
- Zhang, S.; Knight, T.J.; Reecy, J.M.; Wheeler, T.L.; Shackelford, S.D.; Cundiff, L.V.; Beitz, D.C. Associations of polymorphisms in the promoter I of bovine acetyl-CoA carboxylase-alpha gene with beef fatty acid composition. Anim. Genet. 2010, 41, 417–420. [Google Scholar] [CrossRef]
- Matsumoto, H.; Sasaki, K.; Bessho, T.; Kobayashi, E.; Abe, T.; Sasazaki, S.; Oyama, K.; Mannen, H. The SNPs in the ACACA gene are effective on fatty acid composition in Holstein milk. Mol. Biol. Rep. 2012, 39, 8637–8644. [Google Scholar] [CrossRef]
- Wang, X.; Tomso, D.J.; Liu, X.; Bell, D.A. Single nucleotide polymorphism in transcriptional regulatory regions and expression of environmentally responsive genes. Toxicol. Appl. Pharm. 2005, 207, 84–90. [Google Scholar] [CrossRef]
- Badaoui, B.; Serradilla, J.; Tomas, A.; Urrutia, B.; Ares, J.; Carrizosa, J.; Sanchez, A.; Jordana, J.; Amills, M. Goat acetyl-coenzyme A carboxylase α: Molecular characterization, polymorphism, and association with milk traits. J. Dairy Sci. 2007, 90, 1039–1043. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.V.; Jayakumar, S.; Malik, Z.S. Exploring polymorphism and effects of candidate genes on milk fat and protein quality in munjal sheep. Indian J. Small Rumin. 2019, 25, 111–114. [Google Scholar] [CrossRef]
- Kęsek, M.M.; Smołucha, G.; Zielak-Steciwko, A.E. Acetyl-coA cArboxylAse α And steAroyl-coA desAturAse genes polymorphism And their influence on fAtty Acid profile in milk of polish holstein-friesiAn cows. Ann. Anim. Sci. 2017, 17, 993–1006. [Google Scholar] [CrossRef] [Green Version]
- Kęsek-Woźniak, M.M.; Wojtas, E.; Zielak-Steciwko, A.E. Impact of SNPs in ACACA, SCD1, and DGAT1 Genes on Fatty Acid Profile in Bovine Milk with Regard to Lactation Phases. Animals 2020, 10, 997. [Google Scholar] [CrossRef]
- Arnould, V.M.; Soyeurt, H. Genetic variability of milk fatty acids. J. Appl. Genet. 2009, 50, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Liu, C.; Zhang, X.; Liu, X.; Li, W.; Li, F.; Wang, X.; Yue, X. Characterization of fatty acid compositions in longissimus thoracis muscle and identification of candidate gene and SNPs related to polyunsaturated fatty acid in Hu sheep. J. Anim. Sci. 2022, 101, 1–15. [Google Scholar] [CrossRef]
- Henriquez-Rodriguez, E.; Tor, M.; Pena, R.N.; Estany, J. A polymorphism in the stearoyl-CoA desaturase gene promoter increases monounsaturated fatty acid content in dry-cured ham. Meat Sci. 2015, 106, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Ma, Y.; Jiang, L. Review: Research Progress of Dairy Sheep Milk Genes. Agriculture 2022, 12, 169. [Google Scholar] [CrossRef]
- Pegolo, S.; Cecchinato, A.; Mele, M.; Conte, G.; Schiavon, S.; Bittante, G. Effects of candidate gene polymorphisms on the detailed fatty acids profile determined by gas chromatography in bovine milk. J. Dairy Sci. 2016, 99, 4558–4573. [Google Scholar] [CrossRef] [Green Version]
- Dettori, M.L.; Petretto, E.; Mingioni, P.; Vacca, G.M.; Pazzola, M. Variability of genes involved in lipid metabolism and their effect on milk yield, composition and coagulation traits in Sarda sheep. Sci. E Tec. Latt. Casearia 2022, 72, 57–65. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Nutrition | 30% Alfalfa Hay | 70% Concentrate |
|---|---|---|
| Chemical composition (%) | ||
| Dry matter % | 27.41 | 62.97 |
| Crude protein % | 5.19 | 9.11 |
| ME, Mcal/kg | 0.82 | 2.01 |
| NDF % | 13.29 | 26.11 |
| ADF % | 10.97 | 16.87 |
| Ash % | 3.138 | 8.44 |
| Fat % | 0.002 | 1.71 |
| Fatty acid composition (%) | ||
| C6:0 | 1.14 | -- |
| C8:0 | 3.12 | 0.12 |
| C12:0 | 0.48 | -- |
| C14:0 | 1.83 | 0.12 |
| C16:0 | 22.66 | 15.04 |
| C16:1 cis 9 | 1.29 | 0.18 |
| C17:0 | 0.76 | 0.12 |
| C18:0 | 6.26 | 2.29 |
| C18:1 trans 11 | -- | 1.29 |
| C18:1 cis 9 | 10.20 | 23.70 |
| C18:2 cis 9, 12 | 17.42 | 51.43 |
| C20:0 | 3.72 | 0.39 |
| C18:3 cis 9, 12, 15 | 25.32 | 4.93 |
| C22:0 | 3.92 | 0.29 |
| C20:4 cis7, 10, 13, 16 | 1.87 | 0.10 |
| SFA | 43.89 | 18.37 |
| UFA | 56.11 | 81.63 |
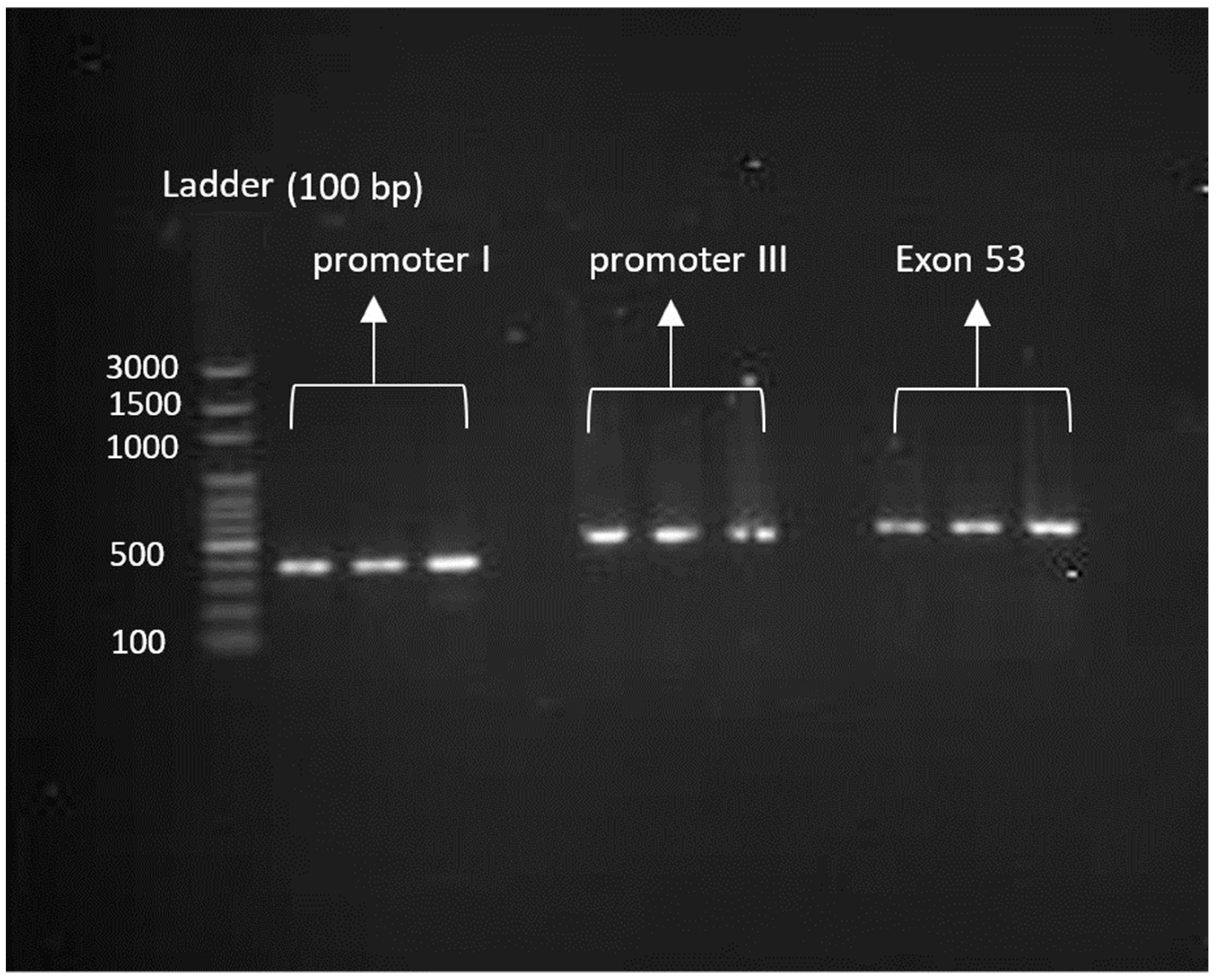
| Target | Sequences | Annealing Temperature (°C) | PCR (bp) |
|---|---|---|---|
| Promoter I | F: GTGGCAAACGTTGTCTTTCT R: CGTATGGGCTTCACTGACTG | 60° | 377 |
| Promoter III | F: GGGCCCCTTTACGTTTCTGT R: AGCTTCTGCCTTAGCTGCA | 61° | 526 |
| Exon 53 | F: CCAGTTATCAGCAGAGGCGG R: GTGGGACTCAGTTTCCCGTC | 60° | 499 |
| SNPs | Acc. No. | Allele Frequencies | Hardy–Weinberg Frequencies | Chi-Squared (χ2) | |||
|---|---|---|---|---|---|---|---|
| PI region | |||||||
| 4412G > A | MT512519 | G = 0.69 | A = 0.31 | GG (36) E = 36.96 | AG (34) E = 32.07 | AA (6) E = 6.96 | 0.27 |
| 4441T > A | MT512520 | T = 0.92 | A = 0.08 | TT (64) E = 64.47 | AT (12) E = 11.05 | AA (0) E = 0.47 | 0.55 |
| 4485C > G | MT512521 | C = 0.34 | G = 0.66 | CC (8) E = 8.89 | CG (36) E = 34.21 | GG (32) E = 32.89 | 0.21 |
| 4507T > C | MT512522 | T = 0.79 | C = 0.21 | TT (42) E = 40.97 | TC (20) E = 22.06 | CC (14) E = 2.97 | 0.57 |
| PIII region | |||||||
| 1007T > G | MT512524 | T = 0.86 | G = 0.14 | TT (57) E = 56.45 | GT (15) E = 18.09 | GG (3) E = 1.45 | 2.15 |
| 1014C > T | MT512525 | C = 0.86 | T = 0.14 | CC (59) E = 56.45 | CT (11) E = 11.95 | TT (5) E = 1.45 | 11.69 |
| 1168A > G | MT512526 | A = 0.43 | G = 0.57 | AA (11) E = 13.90 | AG (42) E = 37.20 | GG (22) E = 24.89 | 1.84 |
| 1331G > T | MT512527 | G = 0.41 | T = 0.59 | GG (10) E = 12.64 | TG (41) E = 36.71 | TT (24) E = 26.64 | 1.57 |
| 1339C > G | MT512528 | C = 0.58 | G = 0.42 | CC (27) E = 25.47 | GC (33) E = 37.05 | GG (15) E = 13.47 | 0.51 |
| 1431C > T | MT512529 | C = 0.81 | T = 0.19 | CC (50) E = 49.76 | TC (21) E = 23.46 | TT (4) E = 2.76 | 0.84 |
| SNPs | Acc. No. | Allele Frequencies | Hardy–Weinberg Frequencies | Chi-Squared (χ2) | |||
|---|---|---|---|---|---|---|---|
| Exon 53 region | |||||||
| 6627G > C | MT649197 | G = 0.69 | C = 0.31 | GG (38) E = 36.26 | CG, (29) E = 32.46 | CC (9) E = 7.26 | 0.86 |
| 6668C < T | MT649197 | C = 0.79 | T = 0.21 | CC (46) E = 45.03 | TC (25) E = 26.94 | TT (5) E = 4.03 | 0.39 |
| 6855T > C | MT649198 | T = 0.74 | C = 0.26 | TT (45) E = 41.26 | TC (22) E = 29.47 | CC (9) E = 5.26 | 4.88 |
| 6860G > C | MT649199 | G = 0.76 | C = 0.24 | GG (49) E = 44.26 | CG (18) E = 27.47 | CC (9) E = 4.26 | 9.04 |
| 6894T > C | MT649200 | T = 0.76 | C = 0.24 | TT (48) E = 44.26 | TC (20) E = 27.47 | CC (8) E = 4.26 | 5.62 |
| 6898C > T | MT649201 | C = 0.80 | T = 0.20 | CC (53) E = 48.16 | TC (15) E = 24.67 | TT (8) E = 3.16 | 11.68 |
| 6977C > G | MT649202 | C = 0.73 | G = 0.27 | CC (44) E = 40.52 | CG (23) E = 29.94 | GG (9) E = 5.53 | 4.08 |
| 6989T > C | MT649203 | T = 0.74 | C = 0.26 | TT (44) E = 41.26 | TC (24) E = 29.47 | CC (8) E = 5.26 | 2.62 |
| 7029G > T | MT649204 | G = 0.68 | T = 0.32 | GG (38) E = 34.89 | GT (27) E = 33.20 | TT (11) E = 7.89 | 2.65 |
| 7031C > T | MT649205 | C = 0.77 | T = 0.23 | CC (49) E = 45.02 | TC (19) E = 26.94 | TT (8) E = 4.02 | 6.60 |
| Milk Traits | Genotypes (Mean ± SE) | p Value | ||
|---|---|---|---|---|
| Promoter I | ||||
| Locus SNP: 4412G > A | GG | AG | AA | |
| C16:0 | 27.33 ± 0.56 a | 26.68 ± 0.57 b | 24.35 ± 1.21 c | 0.04 |
| C16:1 cis7 | 0.29 ± 0.01 c | 0.30 ± 0.01 b | 0.34 ± 0.02 a | 0.03 |
| C18:2 cis9, 12 (LA) | 3.91 ± 0.19 c | 4.01 ± 0.19 b | 4.80 ± 0.32 a | 0.01 |
| Locus SNP: 4441T > A | TT | TA | -- | |
| C22:0 | 0.13 ± 0.02 b | 0.21 ± 0.03 a | -- | 0.01 |
| Locus SNP: 4485C > G | CC | CG | GG | |
| C18:1cis11 (VA) | 0.53 ± 0.02 a | 0.49 ± 0.02 b | 0.48 ± 0.02 c | 0.05 |
| C18:2 (CLA) | 0.82 ± 0.03 a | 0.71 ± 0.03 c | 0.76 ± 0.03 b | 0.03 |
| Locus SNP: 4507C > T | CC | TC | TT | |
| C20:0 | 0.23 ± 0.02 c | 0.32 ± 0.01 a | 0.30 ±0.01b | 0.01 |
| Promoter III | ||||
| Locus SNP: 1007T > G | GG | GT | TT | |
| C12:0 | 4.22 ± 0.62 a | 2.83 ± 0.33 c | 3.19 ± 0.28 b | 0.04 |
| CLA | 0.49 ± 0.10 b | 0.77 ± 0.04 a | 0.76 ± 0.02 ab | 0.03 |
| Locus SNP: 1014C > T | CC | CT | TT | |
| CLA | 0.75 ± 0.02 b | 0.82 ± 0.04 a | 0.57 ± 0.07 c | 0.02 |
| Locus SNP: 1168A > G | AA | AG | GG | |
| Protein | 4.08 ± 0.25 b | 4.58 ± 0.21 a | 4.51 ± 00.21 ab | 0.04 |
| C16:1 cis7 | 0.28 ± 0.01 c | 0.31 ± 0.01 a | 0.29 ± 0.01 b | 0.005 |
| Locus SNP: 1331G > T | GG | GT | TT | |
| Protein | 4.07 ± 0.25 b | 4.56 ± 0.20 a | 4.54 ± 0.20 a | 0.05 |
| C16:1 cis7 | 0.27 ± 0.01 c | 0.31 ± 0.01 a | 0.30 ± 0.01 b | 0.004 |
| Locus SNP:1431C > T | CC | TC | TT | |
| CLA | 0.76 ± 0.03 ab | 0.77 ± 0.03 a | 0.56 ± 0.08 b | 0.05 |
| C20:4 | 0.31 ± 0.02 b | 0.35 ± 0.02 a | 0.27 ± 0.04 c | 0.03 |
| Traits | Genotypes (Mean ± SE) | p Value | ||
|---|---|---|---|---|
| Locus SNP: 6627G > C | GG | CG | CC | |
| C18:2 cis9, 12 (LA) | 4.11 ± 0.16 a | 3.89 ± 0.17 b | 3.49 ± 0.27 c | 0.05 |
| C20:0 | 0.31 ± 0.01 b | 0.28 ± 0.01 c | 0.35 ± 0.02 a | 0.001 |
| C22:0 | 0.14 ± 0.01 b | 0.12 ± 0.01 c | 0.15 ± 0.01 a | 0.04 |
| C22:5 | 0.18 ± 0.01 a | 0.15 ± 0.01 c | 0.16 ± 0.02 b | 0.01 |
| Locus SNP: 6855C > T | CC | TC | TT | |
| C16:1 cis7 | 0.32 ± 0.01 a | 0.28 ± 0.01 c | 0.30 ± 0.01 b | 0.03 |
| C18:2 cis9, 12 (LA) | 3.50 ± 0.26 c | 3.79 ± 0.18 b | 4.11 ± 0.15 a | 0.02 |
| C20:0 | 0.35 ± 0.02 a | 0.28 ± 0.01 c | 0.30 ± 0.01 b | 0.01 |
| Locus SNP: 6860G > C | GG | GC | CC | |
| Fat | 4.16 ± 0.22 a | 3.04 ± 0.40 c | 3.19 ± 0.58 b | 0.03 |
| C20:0 | 0.30 ± 0.01 b | 0.29 ± 0.01 c | 0.35 ± 0.02 a | 0.04 |
| C22:0 | 0.13 ± 0.01 b | 0.12 ± 0.01 c | 0.18 ± 0.01 a | 0.001 |
| Locus SNP: 6894T > C | TT | CT | CC | |
| C18:2 cis9, 12 (LA) | 4.09 ± 0.14 a | 3.82 ± 0.18 b | 3.25 ± 0.28 c | 0.01 |
| C20:0 | 0.31 ± 0.01 b | 0.27 ± 0.01 c | 0.37 ± 0.02 a | 0.001 |
| C18:3 (ALA) | 0.84 ± 0.06 a | 0.75 ± 0.07 b | 0.68 ± 0.09 c | 0.05 |
| C21:0 | 0.08 ± 0.01 b | 0.07 ± 0.01 c | 0.09 ± 0.01 a | 0.03 |
| C22:0 | 0.14 ± 0.01 b | 0.11 ± 0.01 c | 0.15 ± 0.02 a | 0.01 |
| Locus SNP: 6898T > C | TT | TC | CC | |
| C18:2 cis9, 12 (LA) | 3.28 ± 0.28 c | 3.74 ± 0.21 b | 4.08 ± 0.14 a | 0.01 |
| C20:0 | 0.38 ± 0.02 a | 0.28 ± 0.01 c | 0.30 ± 0.01 b | 0.006 |
| C18:3 (ALA) | 0.68 ± 0.09 c | 0.73 ± 0.08 b | 0.84 ± 0.06 a | 0.04 |
| Locus SNP: 6977C > G | CC | CG | GG | |
| C16:1 cis7 | 0.30 ± 0.01 b | 0.28 ± 0.01 c | 0.32 ± 0.01 a | 0.03 |
| C18:2 cis9, 12 (LA) | 4.13 ± 0.15 a | 3.78 ± 0.17 b | 3.50 ± 0.26 c | 0.01 |
| C20:0 | 0.30 ± 0.01 b | 0.28 ± 0.01 c | 0.35 ± 0.02 a | 0.01 |
| Locus SNP: 6989C > T | CC | CT | TT | |
| C18:2 cis9, 12 (LA) | 3.26 ± 0.28 c | 3.95 ± 0.17 b | 4.06 ± 0.14 a | 0.02 |
| C20:0 | 0.37 ± 0.02 a | 0.28 ± 0.01 c | 0.30 ± 0.01 b | 0.004 |
| Locus SNP: 7029G > T | GG | GT | TT | |
| C16:0 | 29.3 ± 0.50 a | 28.1 ± 0.58 b | 26.6 ± 0.91 c | 0.03 |
| C20:0 | 0.30 ± 0.01 b | 0.29 ± 0.01 c | 0.34 ± 0.02 a | 0.03 |
| C21:0 | 0.08 ± 0.01 b | 0.07 ± 0.01 c | 0.10 ± 0.01 a | 0.05 |
| C22:0 | 0.13 ± 0.01 b | 0.12 ± 0.01 c | 0.17 ± 0.01 a | 0.001 |
| Locus SNP: 7031C > T | CC | TC | TT | |
| C18:2 cis9, 12 (LA) | 4.08 ± 0.15 a | 3.86 ± 0.19 b | 3.28 ± 0.28 c | 0.01 |
| C20:0 | 0.29 ± 0.01 c | 0.30 ± 0.01 b | 0.37 ± 0.02 a | 0.01 |
| Trait | HB | SNPs | Position | SE | r2 | T | p Value |
|---|---|---|---|---|---|---|---|
| C16:1 cis7 | H1B1 | PIII- T > G | 1007 | 0.009 | 0.061 | 2.16 | 0.03 |
| H4B1 | PIII- G > T | 1331 | 0.009 | 0.064 | 2.20 | 0.03 | |
| C22:0 | H2B1 | PIII- C > T | 1014 | 0.031 | 0.089 | 2.59 | 0.01 |
| C6:0 | H1B2 | PI- G > A | 4412 | 0.079 | 0.052 | 2.00 | 0.04 |
| H2B2 | PI- T > A | 4441 | 0.078 | 0.053 | 2.03 | 0.04 | |
| C8:0 | H1B2 | PI- G > A | 4412 | 0.079 | 0.052 | 2.00 | 0.04 |
| H2B2 | PI- T > A | 4441 | 0.078 | 0.053 | 2.03 | 0.04 | |
| ALA (n3) | H1B2 | PI- G > A | 4412 | 0.041 | 0.092 | 2.72 | 0.01 |
| H2B2 | PI- T > A | 4441 | 0.040 | 0.084 | 2.59 | 0.01 | |
| H5B3 | E53- C > T | 6898 | 0.043 | 0.098 | 2.73 | 0.01 | |
| H9B3 | E53- C > T | 7031 | 0.043 | 0.062 | 2.12 | 0.03 | |
| Fat % | H3B3 | E53- G > C | 6860 | 0.27 | 0.07 | 2.31 | 0.03 |
| LA (n6) | H1B3 | E53- G > C | 6627 | 0.121 | 0.069 | 2.25 | 0.03 |
| H2B3 | E53- C > T | 6855 | 0.117 | 0.102 | 2.79 | 0.007 | |
| H4B3 | E53- T > C | 6894 | 0.122 | 0.108 | 2.87 | 0.005 | |
| H5B3 | E53- C > T | 6898 | 0.124 | 0.104 | 2.81 | 0.006 | |
| H6B3 | E53- C > G | 6977 | 0.117 | 0.109 | 2.89 | 0.005 | |
| H7B3 | E53- T > C | 6989 | 0.123 | 0.080 | 2.43 | 0.01 | |
| H9B3 | E53- C > T | 7031 | 0.123 | 0.091 | 2.61 | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matar, A.M.; Alharthi, A.S.; Ayadi, M.; Al-Garadi, M.A.; Aljummah, R.S. Identification of Genetic Polymorphisms of PI, PIII, and Exon 53 in the Acetyl-CoA Carboxylase-α (ACACα) Gene and Their Association with Milk Composition Traits of Najdi Sheep. Animals 2023, 13, 1317. https://doi.org/10.3390/ani13081317
Matar AM, Alharthi AS, Ayadi M, Al-Garadi MA, Aljummah RS. Identification of Genetic Polymorphisms of PI, PIII, and Exon 53 in the Acetyl-CoA Carboxylase-α (ACACα) Gene and Their Association with Milk Composition Traits of Najdi Sheep. Animals. 2023; 13(8):1317. https://doi.org/10.3390/ani13081317
Chicago/Turabian StyleMatar, Abdulkareem M., Abdulrahman S. Alharthi, Moez Ayadi, Maged A. Al-Garadi, and Riyadh S. Aljummah. 2023. "Identification of Genetic Polymorphisms of PI, PIII, and Exon 53 in the Acetyl-CoA Carboxylase-α (ACACα) Gene and Their Association with Milk Composition Traits of Najdi Sheep" Animals 13, no. 8: 1317. https://doi.org/10.3390/ani13081317
APA StyleMatar, A. M., Alharthi, A. S., Ayadi, M., Al-Garadi, M. A., & Aljummah, R. S. (2023). Identification of Genetic Polymorphisms of PI, PIII, and Exon 53 in the Acetyl-CoA Carboxylase-α (ACACα) Gene and Their Association with Milk Composition Traits of Najdi Sheep. Animals, 13(8), 1317. https://doi.org/10.3390/ani13081317