


Effect of Sequential Fermentation with Lachancea thermotolerans/S. cerevisiae on Aromatic and Flavonoid Profiles of Plavac Mali Wine




Abstract
:1. Introduction
2. Materials and Methods
2.1. Environmental Conditions and Vineyard Design
2.2. Fermentation Trials
Yeast Species
2.3. Analysis of Standard Components of Wine
2.4. Analysis of Volatile Compounds by GCMS
2.5. Analysis of Polyphenolic Compounds by HPLC
2.6. Statistical Analyses
3. Results and Discussion
3.1. Physiochemical Composition of the Wines Produced by Sequential L, Conventional S and Spontaneous N Fermentation
3.2. Aroma Profile of the Plavac Mali Wines Produced by Sequential L, Conventional S and Spontaneous N Fermentation
3.2.1. Varietal Aroma Compounds in the Plavac Mali Wines Produced by Sequential L, Conventional S and Spontaneous N Fermentation
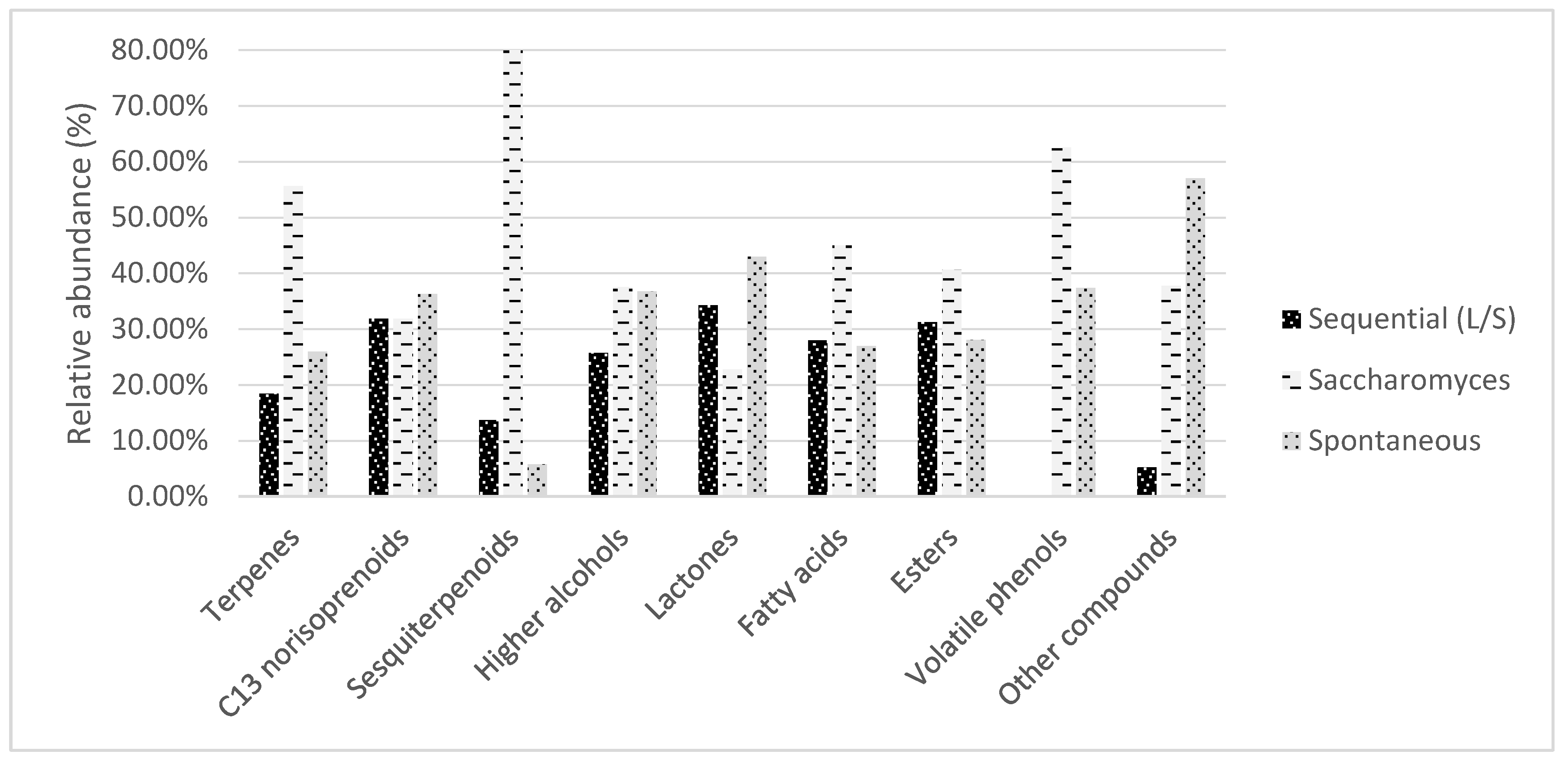
3.2.2. Fermentation Aroma Compounds in the Plavac Mali Wines Produced by Sequential L, Conventional S and Spontaneous N Fermentation
3.3. Polyphenolic Profile of the Plavac Mali Wines Produced by Sequential L, Conventional S and Spontaneous N Fermentation
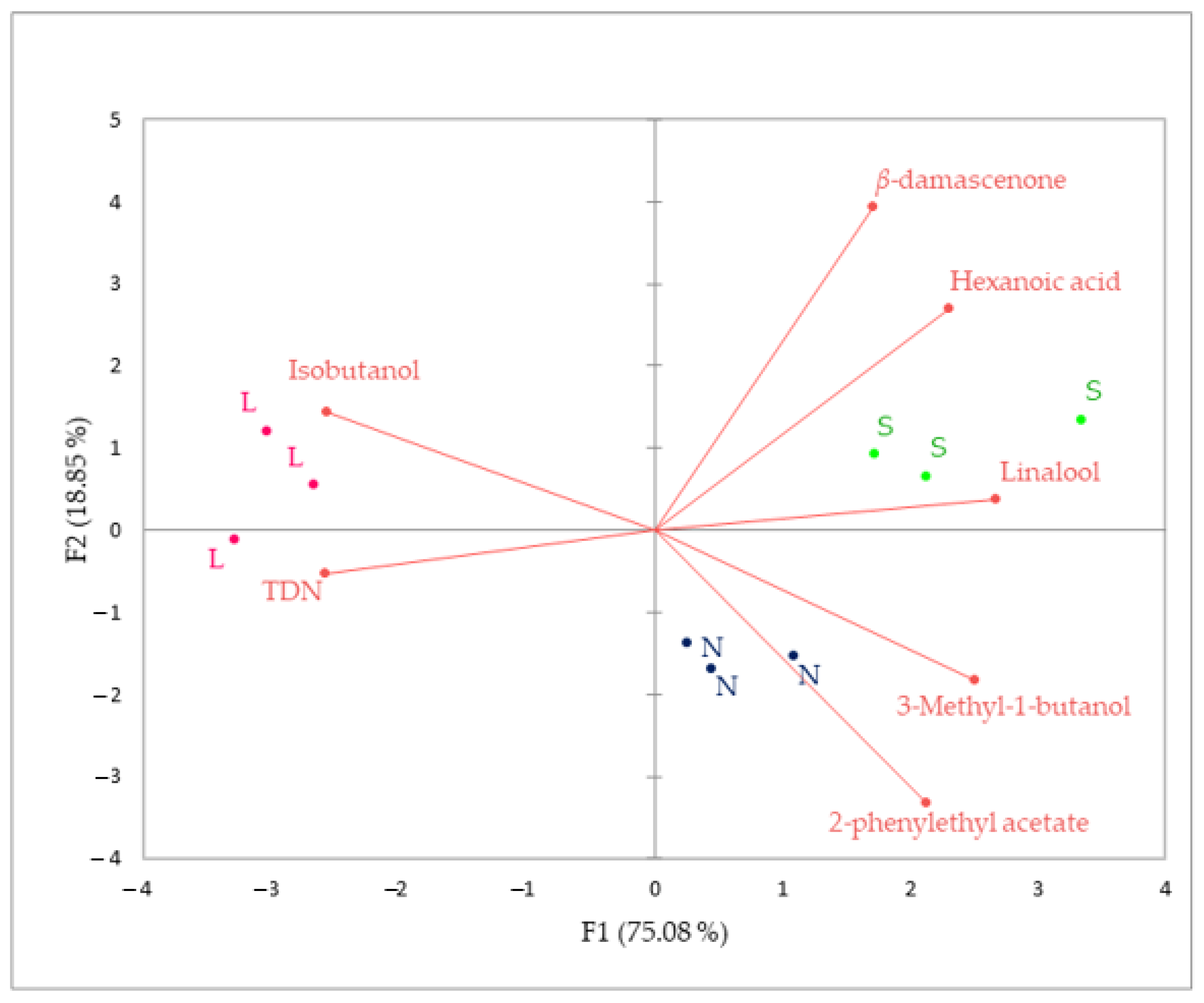
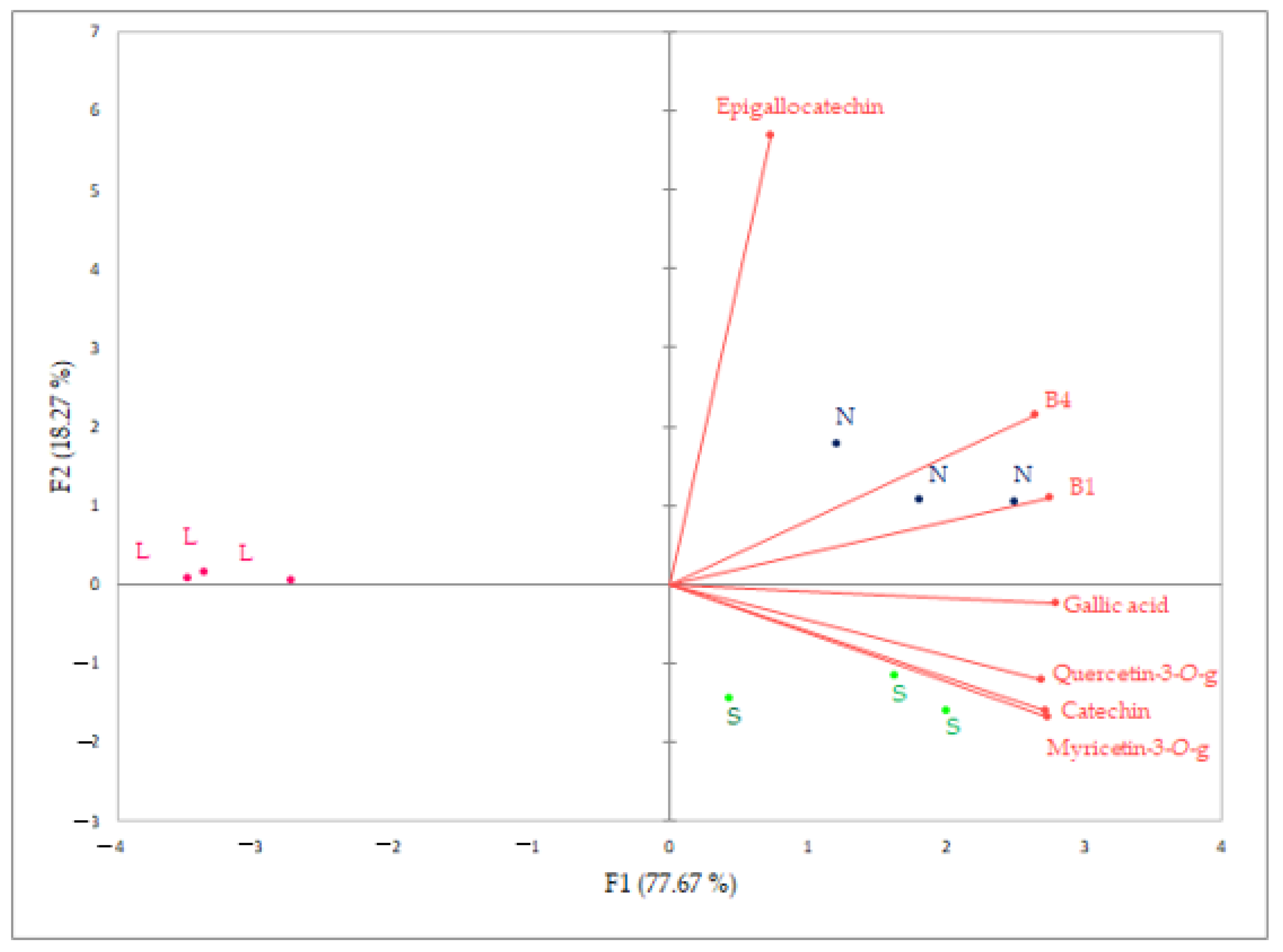
3.4. PCA Analysis of Aroma and Polyphenolic Plavac Mali Wine Composition Produced by Sequential L, Conventional S and Spontaneous N Fermentation
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Capozzi, V.; Garofalo, C.; Chiriatti, M.A.; Grieco, F.; Spano, G. Microbial terroir and food innovation: The case of yeast biodiversity in wine. Microbiol. Res. 2015, 181, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Thorngate, J.H.; Richardson, P.M.; Mills, D.A. Microbial biogeography of wine grapes is conditioned by cultivar, vintage, and climate. Proc. Natl. Acad. Sci. USA 2014, 111, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Fleet, H.G. Wine yeasts for the future. FEMS Yeast Res. 2008, 8, 979–995. [Google Scholar] [CrossRef]
- Han, X.; Xin, Q.; Yang, S.; Li, R.; Huang, W. Study on the diversity of non-Saccharomyces yeasts in Chinese wine regions and their potential in improving wine aroma by β-glucosidase activity analyses. Food Chem. 2021, 360, 129886. [Google Scholar] [CrossRef] [PubMed]
- Martins, G.; Vallance, J.; Mercier, A.; Albertin, W.; Stamatopoulos, P.; Rey, P.; Lonvaud, A.; Masneuf-Pomarède, I. Influence of the farming system on the epiphytic yeasts and yeast-like fungi colonizing grape berries during the ripening process. Int. J. Food Microbiol. 2014, 177, 21–28. [Google Scholar] [CrossRef]
- Shekhawat, K.; Porter, T.J.; Bauer, F.F.; Setati, M.E. Employing oxygen pulses to modulate Lachancea thermotolerans–Saccharomyces cerevisiae Chardonnay fermentations. Ann. Microbiol. 2018, 68, 93–102. [Google Scholar] [CrossRef]
- Hranilovic, A.; Li, S.; Boss, P.K.; Bindon, K.; Ristic, R.; Grbin, P.R.; Jiranek, V. Chemical and sensory profiling of Shiraz wines co-fermented with commercial non-Saccharomyces inocula. Aust. J. Grape Wine Res. 2018, 24, 166–180. [Google Scholar] [CrossRef]
- Molina, A.M.; Swiegers, J.H.; Varela, C.; Pretorius, I.S.; Agosin, E. Influence of wine fermentation temperature on the synthesis of yeast-derived volatile aroma compounds. Appl. Microbiol. Biotechnol. 2007, 77, 675–687. [Google Scholar] [CrossRef]
- Varela, C.; Siebert, T.; Cozzolino, D.; Rose, L.; McLean, H.; Henschke, P.A. Discovering a chemical basis for differentiating wines made by fermentation with ‘wild’ indigenous and inoculated yeasts: Role of yeast volatile compounds. Aust. J. Grape Wine Res. 2009, 15, 238–248. [Google Scholar] [CrossRef]
- Chatonnet, P.; Dubourdieu, D.; Boidron, J.N.; Lavigne, V. Synthesis of volatile phenols by Saccharomyces cerevisiae in wines. J. Sci. Food Agric. 1993, 62, 191–202. [Google Scholar] [CrossRef]
- Hranilovic, A.; Albertin, W.; Capone, D.L.; Gallo, A.; Grbin, P.R.; Danner, L.; Bastian, S.E.; Masneuf-Pomarede, I.; Coulon, J.; Bely, M.; et al. Impact of Lachancea thermotolerans on chemical composition and sensory profiles of Merlot wines. Food Chem. 2021, 349, 129015. [Google Scholar] [CrossRef] [PubMed]
- Jagatić Korenika, A.-M.J.; Tomaz, I.; Preiner, D.; Lavrić, M.; Šimić, B.; Jeromel, A. Influence of L. thermotolerans and S. cerevisiae commercial yeast sequential inoculation on aroma composition of red wines (cv Trnjak, Babic, Blatina and Frankovka). Fermentation 2021, 7, 4. [Google Scholar] [CrossRef]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microb. 2013, 33, 271–281. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; Portu, J.; Garijo, P.; López, R.; Santamaría, P.; López-Alfaro, I.; Gutiérrez, A.R.; González-Arenzana, L. Effect of the sequential inoculation of non-Saccharomyces/Saccharomyces on the anthocyans and stilbenes composition of Tempranillo wines. Front. Microbiol. 2019, 10, 773. [Google Scholar] [CrossRef] [PubMed]
- Benucci, I.; Cerreti, M.; Liburdi, K.; Nardi, T.; Vagnoli, P.; Ortiz-Julien, A.; Esti, M. Pre-fermentative cold maceration in presence of non-Saccharomyces strains: Evolution of chromatic characteristics of Sangiovese red wine elaborated by sequential inoculation. Food Res. Int. 2018, 107, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Porter, T.J.; Divol, B.; Setati, M.E. Investigating the biochemical and fermentation attributes of Lachancea species and strains: Deciphering the potential contribution to wine chemical composition. Int. J. Food Microbiol. 2019, 290, 273–287. [Google Scholar] [CrossRef]
- Gunata, Y.Z.; Bitteur, S.; Brillouet, J.M.; Bayonove, C.L.; Cordonnier, R.E. Sequential enzymic hydrolysis of potentially aromatic glycosides from grape. Carbohyd. Res. 1988, 184, 139–149. [Google Scholar] [CrossRef]
- Escott, C.; Del Fresno, J.M.; Loira, I.; Morata, A.; Tesfaye, W.; del Carmen González, C.; Suárez-Lepe, J.A. Formation of polymeric pigments in red wines through sequential fermentation of flavanol-enriched musts with non-Saccharomyces yeasts. Food Chem. 2018, 239, 975–983. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Benito, S. The combined use of Schizosaccharomyces pombe and Lachancea thermotolerans—Effect on the anthocyanin wine composition. Molecules 2017, 22, 739. [Google Scholar] [CrossRef]
- Casassa, L.F.; Beaver, C.W.; Mireles, M.; Larsen, R.C.; Hopfer, H.; Heymann, H.; Harbertson, J.F. Influence of fruit maturity, maceration length, and ethanol amount on chemical and sensory properties of Merlot wines. Am. J. Enol. Vitic. 2013, 64, 437–449. [Google Scholar] [CrossRef]
- Kontoudakis, N.; Esteruelas, M.; Fort, F.; Canals, J.M.; Zamora, F. Use of unripe grapes harvested during cluster thinning as a method for reducing alcohol content and pH of wine. Aust. J. Grape Wine Res. 2011, 17, 230–238. [Google Scholar] [CrossRef]
- Van Leeuwen, C.; Seguin, G. The concept of terroir in viticulture. J. Wine Res. 2006, 17, 1–10. [Google Scholar] [CrossRef]
- Jagatić Korenika, A.-M.; Tomaz, I.; Preiner, D.; Plichta, V.; Jeromel, A. Impact of commercial yeasts on phenolic profile of Plavac Mali wines from Croatia. Fermentation 2021, 7, 92. [Google Scholar] [CrossRef]
- Winkler, A.J.; Cook, J.A.; Kliewer, W.M.; Lider, L.A. General Viticulture; University of California Press: Berkeley, CA, USA, 1974. [Google Scholar]
- Karoglan, M.; Prtenjak, M.T.; Šimon, S.; Osrečak, M.; Anić, M.; Kontić, J.K.; Andabaka, Ž.; Tomaz, I.; Grisogono, B.; Belušić, A.; et al. Classification of Croatian winegrowing regions based on bioclimatic indices. Paper presented at the E3S Web of Conferences, XIIth International Terroir Congress, Zaragoza, Spain, 18–22 June 2018; Volume 50, p. 01032. [Google Scholar] [CrossRef]
- OIV. Compendium of International Methods of Wine and Must Analysis; International, Organisation of Vine and Wine: Paris, France, 2016. [Google Scholar]
- Šikuten, I.; Štambuk, P.; Karoglan Kontić, J.; Maletić, E.; Tomaz, I.; Preiner, D. Optimization of SPME-Arrow-GC/MS method for determination of free and bound volatile organic compounds from grape skins. Molecules 2021, 26, 7409. [Google Scholar] [CrossRef] [PubMed]
- Song, N.E.; Lee, J.Y.; Lee, Y.Y.; Park, J.D.; Jang, H.W. Comparison of headspace-SPME and SPME-Arrow-GC-MS methods for the determination of volatile compounds in Korean salt-fermented fish sauce. Appl. Biol. Chem. 2019, 62, 16. [Google Scholar] [CrossRef]
- Tomaz, I.; Maslov, L. Simultaneous determination of phenolic compounds in different matrices using phenyl-hexyl stationary phase. Food Anal. Methods 2016, 9, 401–410. [Google Scholar] [CrossRef]
- Berente, B.; De La Calle García, D.; Reichenbächer, M.; Danzer, K. Method development for the determination of anthocyanins in red wines by high-performance liquid chromatography and classification of German red wines by means of multivariate statistical methods. J. Chromatogr. A 2000, 871, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Genovese, A.; Dimaggio, R.; Lisanti, M.T.; Piombino, P.; Moio, L. Aroma composition of red wines by different extraction methods and gas chromatography-SIM/MASS spectrometry analysis. Ann. Chim. 2005, 95, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Loscos, N.; Hernandez-Orte, P.; Cacho, J.; Ferreira, V. Release and formation of varietal aroma compounds during alcoholic fermentation from nonfloral grape odorless flavor precursors fractions. J. Agric. Food Chem. 2007, 55, 6674–6684. [Google Scholar] [CrossRef]
- Genovese, A.; Caporaso, N.; Moio, L. Influence of yeast strain on odor-active compounds in Fiano wine. Appl. Sci. 2021, 11, 7767. [Google Scholar] [CrossRef]
- Carrau, F.M.; Medina, K.; Boido, E.; Farina, L.; Gaggero, C.; Dellacassa, E.; Versini, G.; Henschke, P.A. De novo synthesis of monoterpenes by Saccharomyces cerevisiae wine yeasts. FEMS Microbiol. Lett. 2005, 243, 107–115. [Google Scholar] [CrossRef]
- Keyzers, R.A.; Boss, P.K. Changes in the volatile compound production of fermentations made from musts with increasing grape content. J. Agric. Food Chem. 2010, 58, 1153–1164. [Google Scholar] [CrossRef]
- Cameleyre, M.; Lytra, G.; Tempere, S.; Barbe, J.C. Olfactory impact of higher alcohols on red wine fruity ester aroma expression in model solution. J. Agric. Food Chem. 2015, 63, 9777–9788. [Google Scholar] [CrossRef] [PubMed]
- De-La-Fuente-Blanco, A.; Sáenz-Navajas, M.P.; Ferreira, V. On the effects of higher alcohols on red wine aroma. Food Chem. 2016, 210, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Guth, H. Quantitation and sensory studies of character impact odorants of different white wine varieties. J. Agric. Food Chem 1997, 45, 3027–3032. [Google Scholar] [CrossRef]
- Nan, L.; Liu, L.; Li, Y.; Huang, J.; Wang, Y.; Wang, C.; Wang, Z.; Xu, C. Comparison of aroma compounds in Cabernet Sauvignon red wines from five growing regions in Xinjiang in China. J. Food Qual. 2021, 2021, 5562518. [Google Scholar] [CrossRef]
- Balikci, E.K.; Tanguler, H.; Jolly, N.P.; Erten, H. Influence of Lachancea thermotolerans on cv. Emir wine fermentation. Yeast 2016, 33, 313–321. [Google Scholar] [CrossRef]
- Delgado, J.A.; Sánchez-Palomo, E.; Alises, M.O.; Viñas, M.G. Chemical and sensory aroma typicity of La Mancha Petit Verdot wines. LWT 2022, 162, 113418. [Google Scholar] [CrossRef]
- Romano, P.; Braschi, G.; Siesto, G.; Patrignani, F.; Lanciotti, R. Role of yeasts on the sensory component of wines. Foods 2022, 11, 1921. [Google Scholar] [CrossRef]
- Barbosa, C.; Mendes-Faia, A.; Mendes-Ferreira, A. The nitrogen source impacts major volatile compounds released by Saccharomyces cerevisiae during alcoholic fermentation. Int. J. Food Microbiol. 2012, 160, 87–93. [Google Scholar] [CrossRef]
- Qian, X.; Liu, Y.; Zhang, G.; Yan, A.; Wang, H.; Wang, X.; Pan, Q.; Xu, H.; Sun, L.; Zhu, B. Alcohol acyltransferase gene and ester precursors differentiate composition of volatile esters in three interspecific hybrids of Vitis labrusca× V. vinifera during berry development period. Food Chem. 2019, 295, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Fairbairn, S.; McKinnon, A.; Musarurwa, H.T.; Ferreira, A.C.; Bauer, F.F. The impact of single amino acids on growth and volatile aroma production by Saccharomyces cerevisiae strains. Front. Microbiol. 2017, 8, 2554. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Sánchez, F.J.; Páez-Lerma, J.B.; Rojas-Contreras, J.A.; López-Miranda, J.; Soto-Cruz, N.Ó.; Reinhart-Kirchmayr, M. Study of the enzymatic capacity of Kluyveromyces marxianus for the synthesis of esters. Microb. Physiol. 2019, 29, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mendes-Ferreira, A.; Barbosa, C.; Falco, V.; Leão, C.; Mendes-Faia, A. The production of hydrogen sulphide and other aroma compounds by wine strains of Saccharomyces cerevisiae in synthetic media with different nitrogen concentrations. JIMB 2009, 36, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Ruiz, A.; Alonso-del-Real, J.; Barrio, E.; Querol, A. Saccharomyces cerevisiae wine strains show a wide range of competitive abilities and differential nutrient uptake behavior in co-culture with S. kudriavzevii. Food Microbiol. 2023, 114, 104276. [Google Scholar] [CrossRef]
- Ferreira, V.; López, R.; Cacho, J.F. Quantitative determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agric. 2000, 80, 1659–1667. [Google Scholar] [CrossRef]
- Silva, R.; Coelho, E.; Aguiar, T.Q.; Domingues, L. Microbial biosynthesis of lactones: Gaps and opportunities towards sustainable production. Appl. Sci. 2021, 11, 8500. [Google Scholar] [CrossRef]
- Chou, H.-C.; Šuklje, K.; Antalick, G.; Schmidtke, L.M.; Blackman, J.W. Late-season Shiraz berry dehydration that alters composition and sensory traits of wine. J. Agric. Food Chem. 2018, 66, 7750–7757. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; González, C.; Escott, C. Non-Saccharomyces as biotools to control the production of off-flavors in wines. Molecules 2021, 26, 4571. [Google Scholar] [CrossRef] [PubMed]
- Coghe, S.; Benoot, K.; Delvaux, F.; Vanderhaegen, B.; Delvaux, F.R. Ferulic acid release and 4-vinylguaiacol formation during brewing and fermentation: Indications for feruloyl esterase activity in Saccharomyces cerevisiae. J. Agric. Food Chem. 2004, 52, 602–608. [Google Scholar] [CrossRef]
- Weldegergis, B.T.; Crouch, A.M.; Górecki, T.; De Villiers, A. Solid phase extraction in combination with comprehensive two-dimensional gas chromatography coupled to time-of-flight mass spectrometry for the detailed investigation of volatiles in South African red wines. Anal. Chim. Acta. 2011, 701, 98–111. [Google Scholar] [CrossRef]
- Carrasco-Sánchez, V.; John, A.; Marican, A.; Santos, L.S.; Laurie, V.F. Removal of 4-ethylphenol and 4-ethylguaiacol with polyaniline-based compounds in wine-like model solutions and red wine. Molecules 2015, 20, 14312–14325. [Google Scholar] [CrossRef]
- Perestrelo, R.M.D.S.; Fernandes, A.; Albuquerque, F.F.; Marques, J.C.; Câmara, J.D.S. Analytical characterization of the aroma of Tinta Negra Mole red wine: Identification of the main odorants compounds. Anal. Chim. Acta 2006, 563, 154–164. [Google Scholar] [CrossRef]
- Monagas, M.; Bartolomé, B.; Gómez-Cordovés, C. Evolution of polyphenols in red wines from Vitis vinifera L. during aging in the bottle. Eur. Food Res. Technol. 2005, 220, 331–340. [Google Scholar] [CrossRef]
- Razmkhab, S.; Lopez-Toledano, A.; Ortega, J.M.; Mayen, M.; Merida, J.; Medina, M. Adsorption of phenolic compounds and browning products in white wines by yeasts and their cell walls. J. Agric. Food Chem. 2002, 50, 7432–7437. [Google Scholar] [CrossRef] [PubMed]
- De Melo Lopes, L.M.; Costa Batista, L.H.; Gouveia, M.J.; Leite, T.C.C.; de Mello, M.R.F.; de Assis, S.A.; de Sena, A.R. Kinetic and thermodynamic parameters, and partial characterization of the crude extract of tannase produced by Saccharomyces cerevisiae CCMB 520. Nat. Prod. Res. 2018, 32, 1068–1075. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Gallego, R.; Hernández-Hierro, J.M.; Rivas-Gonzalo, J.C.; Escribano-Bailón, M.T. Sensory evaluation of bitterness and astringency sub-qualities of wine phenolic compounds: Synergistic effect and modulation by aromas. Food Res. Int. 2014, 62, 1100–1107. [Google Scholar] [CrossRef]
- Morata, A.; Gómez-Cordovés, M.C.; Calderón, F.; Suárez, J.A. Effects of pH, temperature and SO2 on the formation of pyranoanthocyanins during red wine fermentation with two species of Saccharomyces. Int. J. Food Microbiol. 2006, 106, 123–129. [Google Scholar] [CrossRef]
- Schwarz, M.; Wabnitz, T.C.; Winterhalter, P. Pathway leading to the formation of anthocyanin−vinylphenol adducts and related pigments in red wines. J. Agric. Food Chem. 2003, 51, 3682–3687. [Google Scholar] [CrossRef]
- Cerpa-Calderón, F.K.; Kennedy, J.A. Berry integrity and extraction of skin and seed proanthocyanidins during red wine fermentation. J. Agric. Food Chem. 2008, 56, 9006–9014. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fermentation | L | S | N |
| Yeast species | Lachancea thermotolerans Saccharomyces cerevisiae | Saccharomyces cerevisiae | Spontaneous yeasts |
| Strain | Kt 421 10 | 10 | Unknown |
| Suppliers | Viniflora, Concerto Siha, Active Yeast 10 | Siha, Active Yeast 10 | Natural |
| Timing of yeast inoculation | after destemming after 24 h | after destemming | No |
| Temperature (°C) | 25.8–26.6 | 26.4–26.9 | 24.8–25.6 |
| Frequency of punch down | every 12 h/5 min | every 12 h/5 min | every 12 h/5 min |
| Duration of maceration | 7 days | 7 days | 7 days |
| Compound | L | S | N |
|---|---|---|---|
| Alcohol (% v/v) | 14.93 ± 0.06 b | 15.13 ± 0.06 ab | 15.23 ± 0.12 a |
| Reducing sugar (g/L) | 7.13 ± 0.81 a | 4.73 ± 0.38 b | 5.27 ± 1.27 ab |
| Extract (g/L) | 25.43 ± 0.61 a | 24.77 ± 0.95 a | 25.43 ± 0.23 a |
| Ash (g/L) | 2.81 ± 0.22 a | 3.11 ± 0.01 a | 2.91 ± 0.05 a |
| pH | 3.56 ± 0.03 b | 3.59 ± 0.03 b | 3.68 ± 0.02 a |
| Total acidity (g/L as tartaric acid) | 5.90 ± 0.00 a | 5.60 ± 0.00 b | 5.30 ± 0.10 c |
| Volatile acidity (g/L as acetic acid) | 0.50 ± 0.00 a | 0.40 ± 0.00 b | 0.40 ± 0.00 b |
| Aroma Compound (µg/L)/Yeast | L | S | N |
|---|---|---|---|
| α-Ocimene | nd | 32.84 ± 1.33 | nd |
| Citronellol | 134.90 ± 10.47 c | 657.16 ± 13.62 a | 201.63 ± 4.32 b |
| α-Pinene | 1.11 ± 0.12 c | 8.60 ± 0.68 a | 4.61 ± 0.41 b |
| α-terpinene | nd | 6.29 ± 0.67 b | 11.61 ± 0.21 a |
| α-terpineol | 4.78 ± 0.97 c | 39.29 ± 0.92 a | 7.19 ± 0.29 b |
| D-Limonene | nd | 26.84 ± 0.99 a | 17.71 ± 2.03 b |
| Hotrienol | 3.12 ± 0.16 c | 8.95 ± 0.25 a | 7.30 ± 0.17 b |
| Linalool | 10.29 ± 0.30 c | 37.53 ± 0.68 a | 25.88 ± 3.33 b |
| β-myrcene | 107.20 ± 5.32 a | 95.21 ± 2.11 a | 111.06 ± 9.56 a |
| Menthol | 36.62 ± 2.07 b | 43.79 ± 2.81 a | 39.20 ± 0.85 ab |
| Citronellol acetate | 5.38 ± 0.48 | nd | nd |
| trans-linalool oxide (furanoid) | 22.30 ± 0.82 b | 26.55 ± 2.84 ab | 32.65 ± 3.50 a |
| Ʃ Terpenes | 325.71 ± 12.66 c | 983.04 ± 10.15 a | 458.84 ± 15.04 b |
| β-damascenone | 5.81 ± 0.52 ab | 6.68 ± 0.30 a | 5.72 ± 0.14 b |
| TDN | 7.27 ± 0.23 a | 6.50 ± 0.24 b | 6.84 ± 0.12 ab |
| TPB | 3.32 ± 0.28 c | 6.57 ± 0.20 b | 8.91 ± 0.86 a |
| Vitispirane A | 14.50 ± 0.89 b | 22.87 ± 0.93 a | 17.62 ± 1.06 ab |
| Vitispirane B | 11.69 ± 0.78 b | 19.36 ± 2.01 a | 16.73 ± 1.26 a |
| 2,3-dehydro-4-oxo-β-ionol | 12.18 ± 0.49 a | nd | 8.01 ± 0.14 b |
| 4-hydroxy-β-ionone | 5.66 ± 0.40 a | 4.19 ± 0.84 b | 5.45 ± 0.30 ab |
| 2,5,8-trimethyl-1,2,3,4-tetrahydro-1-naphthalenol | 5.85 ± 0.49 a | nd | 6.13 ± 0.18 a |
| Ʃ Norisoprenoids | 66.28 ± 2.83 b | 66.17 ± 2.21 b | 75.41 ± 0.68 a |
| cis-α-bisabolene | 5.45 ± 0.39 | nd | nd |
| Isocaryophyllene | 15.14 ± 0.68 | nd | nd |
| cis-β-farnesene | 18.60 ± 1.13 b | 231.04 ± 20.71 a | 16.60 ± 0.99 b |
| Ʃ Sesquiterpenoids | 39.19 ± 0.87 b | 231.04 ± 20.71 a | 16.6 ± 0.99 b |
| cis-6-nonen-1-ol | 16.57 ± 0.40 b | 17.57 ± 1.90 ab | 20.29 ± 0.91 a |
| 1,3-octanediol | 3.08 ± 0.29 a | nd | 2.76 ± 0.41 a |
| 1-butanol | 1617.69 ± 41.58 b | 1386.23 ± 76.94 b | 2551.74 ± 183.83 a |
| 3-methyl-1-butanol (Isoamyl alc) | 77311.14 ± 1411.88 b | 114386.01 ± 4223.04 a | 115538.37 ± 5480.85 a |
| 1-decanol | 5899.25 ± 113.55 c | 14565.27± 300.42 a | 11731.37 ± 269.77 b |
| 2-ethyl-1-hexanol | 41.21 ± 1.83 a | 34.23 ± 3.93 b | nd |
| 1-hexanol | 8694.68 ± 167.31 b | 13825.33 ± 1033.70 a | 13957.16 ± 344.91 a |
| 1-dodecanol | nd | 8721.94 ± 153.36 a | 6168.52 ± 285.03 b |
| 1-nonanol | 33.43 ± 1.18 c | 60.87 ± 2.26 a | 53.72 ± 1.63 b |
| 1-octanol | 72.78 ± 2.34 b | 158.91 ± 4.90 a | 164.51 ± 3.51 a |
| 1-octen-3-ol | 65.00 ± 1.29 b | 90.10 ± 2.39 a | 94.44 ± 5.72 a |
| 1-pentanol | 3347.18 ± 84.22 b | 4621.36 ± 144.71 a | 4605.15 ± 252.26 a |
| 4-methyl-1-pentanol | 462.46 ± 9.84 c | 1994.52 ± 41.12 b | 2595.56 ± 184.89 a |
| 1-propanol | 5259.57 ± 90.84 a | 805.32 ± 67.88 c | 1110.80 ± 128.61 b |
| 3-ethoxy-1-propanol | 1833.89 ± 11.51 a | 76.67 ± 2.94 b | 30.29 ± 1.09 c |
| 3-ethyl-4-methylpentan-1-ol | 4013.40 ± 89.47 a | 1204.50 ± 173.69 c | 1682.82 ± 58.59 b |
| trans-3-hexen-1-ol | 286.54 ± 10.56 b | 5268.64 ± 132.29 a | 5299.20 ± 286.28 a |
| cis-3-hexen-1-ol | 696.89 ± 13.14 b | 692.10 ± 17.18 b | 985.02 ± 43.83 a |
| 2-heptanol | 7.36 ± 0.46 b | 8.02 ± 0.75 b | 12.67 ± 2.33 a |
| 2-nonanol | 8.27 ± 1.21 a | 9.87 ± 1.21 a | 9.91 ± 0.25 a |
| trans-2-octen-1-ol | 37.62 ± 2.99 a | 41.62 ± 0.77 a | 38.36 ± 1.06 a |
| 3-methylpentan-1-ol | 2008.78 ± 40.89 | nd | nd |
| 3-octanol | 30.45 ± 0.76 b | 30.01 ± 1.76 b | 54.18 ± 1.15 a |
| 3-methyl-3-pentanol | 1221.41 ± 22.51 | nd | nd |
| 3-penten-1-ol | 1816.10 ± 72.50 c | 5797.31 ± 104.23 a | 3722.42 ± 116.91 b |
| Isobutanol | 8618.19 ± 153.22 a | 7359.04 ± 430.46 b | 7477.05 ± 135.44 b |
| 2-phenylethanol | 4639.36 ± 88.67 c | 6299.05 ± 144.66 a | 5868.90 ± 191.30 b |
| 2,3-butanediol | 1197.41 ± 21.02 a | 1233.12 ± 47.03 a | 811.39 ± 14.49 b |
| Ʃ Higher alcohols | 129239.69 ± 1889.85 b | 188687.6 ± 4988.08 a | 184586.61 ± 7231.42 a |
| trans-oak lactone | nd | 16.95 ± 1.15 | nd |
| γ-heptalactone | 183.47 ± 2.62 c | 461.44 ± 11.83 a | 391.96 ± 12.44 b |
| Butyrolactone | 561.67 ± 10.20 a | 17.01 ± 0.62 b | 541.83 ± 21.09 a |
| Ʃ Lactones | 745.14 ± 11.52 b | 495.39 ± 11.85 c | 933.8 ± 32.44 a |
| 2,4-hexadienedioic | 3.00 ± 0.28 b | nd | 4.34 ± 0.59 a |
| 3-methylbutanoic acid | 5090.29 ± 89.57 b | 7670.03 ± 49.48 a | 3719.85 ± 52.49 c |
| Butanoic acid | 768.12 ± 15.11 a | nd | 300.17 ± 7.78 b |
| 4-hydroxy-butanoic acid | nd | 870.72 ± 17.53 | nd |
| Decanoic acid | 359.63 ± 8.29 b | 250.41 ± 49.27 c | 525.00 ± 32.10 a |
| Dodecanoic acid | 22.15 ± 2.25 a | nd | 24.30 ± 1.34 a |
| Hexanoic acid | 1902.41 ± 33.67 b | 3240.13 ± 167.50 a | 2120.80 ± 102.51 b |
| 2-Ethyl hexanoic acid | nd | nd | 81.03 ± 1.52 |
| Nonanoic acid | 67.28 ± 2.53 c | 167.85 ± 4.93 a | 117.54 ± 2.07 b |
| Octanoic acid | 991.91 ± 17.59 c | 2622.32 ± 79.09 a | 1982.15 ± 44.84 b |
| trans-2-undecenoic acid | nd | nd | 10.57 ± 0.79 |
| Ʃ Fatty acids | 9204.78 ± 161.32 b | 14821.45 ± 335.48 a | 8885.76 ± 122.63 b |
| 3-methyl-1-butanoate | 726.90 ± 13.96 b | 2781.87 ± 214.68 a | 974.04 ± 61.19 b |
| Ethyl-2,4-hexadienoate | 4.50 ± 0.52 c | 11.98 ± 1.20 a | 6.94 ± 0.60 b |
| Ethyl-2-hexenoate | 3.35 ± 0.40 b | 19.08 ± 1.30 a | 21.24 ± 1.34 a |
| Diethyl-malate | 68.68 ± 2.31 b | 111.72 ± 10.14 a | 79.49 ± 1.42 b |
| 3-methylbuthyl decanoate | 6.08 ± 0.73 b | nd | 7.45 ± 0.52 a |
| 2-phenylethyl acetate | 642.87 ± 11.88 c | 885.48 ± 19.53 b | 1008.36 ± 15.32 a |
| Hexyl acetate | 19.65 ± 1.32 b | 48.49 ± 2.85 a | 13.16 ± 0.60 c |
| Pentyl acetate | 2.63 ± 0.10 b | nd | 3.30 ± 0.23 a |
| Phenyl-ethyl acetate | 51.89 ± 1.93 b | 80.09 ± 3.83 a | 83.83 ± 4.52 a |
| Methyl-2-hydroxybenzoate | 4.36 ± 0.55 b | 7.83 ± 0.65 a | 2.00 ± 0.24 c |
| Diethyl butanoate | 869.19 ± 17.30 b | 99.75 ± 5.79 c | 978.94 ± 70.75 a |
| Ethyl butanoate | 42.07 ± 1.13 a | nd | 34.27 ± 2.58 b |
| Ethyl decanoate | 95.59 ± 3.88 b | 96.76 ± 4.80 b | 196.49 ± 7.68 a |
| Methyl decanoate | nd | nd | 6.12 ± 0.14 |
| Ethyl dodecanoate | 14.93 ± 0.90 b | 8.74 ± 0.50 c | 18.35 ± 1.24 a |
| Ethyl-2-hydroxy-4-methylpentanoate | 40.65 ± 0.57 b | 76.86 ± 3.65 a | 40.38 ± 1.06 b |
| Ethyl-2-hydroxypropanoate | 622.32 ± 11.40 a | 572.38 ± 15.28 b | 364.21 ± 8.10 c |
| Ethyl-3-hydroxybutanoate | 22.94 ± 0.74 c | 44.27 ± 2.24 b | 55.29 ± 2.00 a |
| Ethyl-4-hydroxybutanoate | 283.38 ± 3.00 c | 434.84 ± 23.11 a | 353.89 ± 15.30 b |
| Ethyl-9-decanoate | 132.58 ± 5.34 a | 126.48 ± 2.72 a | 128.17 ± 2.27 a |
| Ethyl-9-hexadecenoate | 13.28 ± 0.71 b | 23.83 ± 2.41 a | 24.95 ± 1.59 a |
| Ethyl hydrogen succinate | 278.97 ± 11.75 c | 795.38 ± 69.54 a | 522.43 ± 24.67 b |
| Ethyl cinnamate | 64.64 ± 4.11 a | nd | 61.52 ± 1.53 a |
| trans-3-hexen-1-ol acetate | 93.69 ± 3.09 b | 914.22 ± 31.67 a | 9.37 ± 1.26 c |
| cis-3-hexen-1-ol acetate | 1.12 ± 0.34 b | 2.66 ± 0.26 a | nd |
| Ethyl heptanoate | nd | 21.13 ± 0.50 | nd |
| Ethyl hexadecanoate | 38.66 ± 0.68 a | 36.61 ± 0.87 b | 39.98 ± 0.72 a |
| Ethyl-3-hydroxyhexanoate | nd | nd | 3.87 ± 0.12 |
| Isoamyl acetate | nd | 16.22 ± 0.33 a | 0.23 ± 0.06 b |
| Ethyl nonanoate | 52.84 ± 1.34 a | 52.77 ± 1.52 a | 53.29 ± 0.91 a |
| 3-methylbuthyl ocatnoate | 8.35 ± 0.42 b | 14.40 ± 1.69 a | 11.58 ± 1.19 a |
| Ethyl octanoate | 244.15 ± 5.57 b | 779.85 ± 20.57 a | 758.85 ± 30.17 a |
| Methyl octanoate | 17.19 ± 1.60 b | nd | 19.96 ± 0.66 a |
| 3-methylbutyl pentadecanoate | 7.41 ± 0.23 b | 9.16 ± 0.12 a | 9.74 ± 0.43 a |
| Ethyl pentadecanoate | 31.10 ± 0.86 c | 61.59 ± 2.89 a | 40.56 ± 2.47 b |
| 2-methyl-propanoate | 4002.09 ± 92.78 a | 2908.13 ± 120.07 b | 1645.54 ± 27.98 c |
| Butyl ethyl succinate | 55.95 ± 3.69 c | 94.18 ± 4.13 b | 112.44 ± 3.63 a |
| Ʃ Esters | 8563.98 ± 79.48 b | 11136.77 ± 86.63 a | 7690.21 ± 61.69 c |
| 4-ethyl guaiacol | nd | 17.67 ± 1.41 a | 14.38 ± 0.67 b |
| 4-ethyl phenol | nd | 96.27 ± 2.61 | nd |
| Eugenol | nd | nd | 53.76 ± 1.06 |
| Ʃ Volatile phenols | 0 ± 0 | 113.94 ± 2.09 a | 68.13 ± 1 b |
| Benzaldehyde | 58.03 ± 2.23 b | 566.09 ± 18.47 a | 42.10 ± 1.66 b |
| Benzyl alcohol | 422.79 ± 21.06 b | 1610.89 ± 147.13 a | 521.31 ± 15.57 b |
| Furfuryl alcohol | nd | nd | 150.06 ± 10.41 |
| Furfural | nd | 260.99 ± 9.82 | nd |
| 2,4-dimethyl-3-pentanol | 79.26 ± 4.26 b | nd | 112.28 ± 8.88 a |
| 6-methyl-5-hepten-2-one | 3.21 ± 0.07 b | 9.85 ± 0.17 a | 11.93 ± 1.90 a |
| Acetoin | 14.42 ± 1.67 b | nd | 22.24 ± 2.49 a |
| Methionol | 53.23 ± 3.53 c | 2080.22 ± 45.18 b | 5978.68 ± 107.43 a |
| Ʃ Other compounds | 630.95 ± 22.45 c | 4528.04 ± 162.84 b | 6838.61 ± 98.14 a |
| Compound (mg/L) | L | S | N |
|---|---|---|---|
| Caftaric acid | 27.01 ± 1.34 b | 27.98 ± 1.32 b | 35.12 ± 0.59 a |
| Caffeic acid | 9.56 ± 1.15 a | 10.31 ± 0.97 a | 11.74 ± 1.18 a |
| Ʃ HCA | 36.57 ± 2.49 b | 38.28 ± 2.27 b | 46.86 ± 1.41 a |
| Resveratrol 3-O-glucoside | 7.18 ± 0.28 b | 7.80 ± 0.33 ab | 8.08 ± 0.17 a |
| Ʃ Stilbenes | 7.18 ± 0.28 b | 7.80 ± 0.33 ab | 8.08 ± 0.17 a |
| Gallic acid | 31.11 ± 0.25 b | 34.25 ± 1.16 a | 34.52 ± 1.00 a |
| Protocatechuic acid | 3.93 ± 0.05 a | 3.82 ± 0.15 a | 3.89 ± 0.08 a |
| Vanillic acid | 5.43 ± 0.26 a | 5.11 ± 0.37 a | 5.53 ± 0.28 a |
| Syringic acid | 11.42 ± 0.23 b | 13.50 ± 0.31 a | 11.99 ± 0.10 b |
| Ʃ HBA | 51.90 ± 0.61 b | 56.69 ± 1.19 a | 55.93 ± 1.09 a |
| Delphinidin-3-O-glucoside | 3.45 ± 0.17 c | 4.19 ± 0.12 b | 4.69 ± 0.15 a |
| Petunidin-3-O-glucoside | 8.47 ± 0.15 b | 9.80 ± 0.11 a | 10.10 ± 0.33 a |
| Peonidin-3-O-glucoside | 5.63 ± 0.37 b | 7.22 ± 0.14 a | 7.26 ± 0.11 a |
| Malvidin-3-O-glucoside | 70.32 ± 0.69 b | 85.30 ± 2.80 a | 82.05 ± 1.79 a |
| Cyanidin-3-O-glucoside | 0.55 ± 0.21 a | 0.29 ± 0.02 a | 0.54 ± 0.16 a |
| Malvidin-3-(6-O-acetyl) glucoside | 4.69 ± 0.13 b | 5.66 ± 0.29 a | 5.78 ± 0.13 a |
| Malvidin-3-O-glucoside-cafeoil-g | 0.53 ± 0.03 c | 0.73 ± 0.05 b | 0.83 ± 0.02 a |
| Peonidin-3-O-(p-coumaroyl)-glucoside | 0.20 ± 0.06 b | 0.67 ± 0.10 a | 0.64 ± 0.02 a |
| Malvidin-3-O-(p-coumaroyl)-glucoside | 8.78 ± 0.25 b | 10.31 ± 0.20 a | 10.16 ± 0.27 a |
| Ʃ Anthocyanins | 102.61 ± 0.78 b | 124.16 ± 2.73 a | 122.06 ± 2.20 a |
| Myricetin-3-O-glucoside | 4.54 ± 0.22 b | 5.42 ± 0.12 a | 5.44 ± 0.25 a |
| Myricetin-3-O-glucuronide | 0.72 ± 0.07 b | 0.60 ± 0.08 b | 1.02 ± 0.06 a |
| Quercetin-3-O-galactoside | 0.20 ± 0.04 b | 0.32 ± 0.03 a | 0.31 ± 0.01 a |
| Quercetin-3-O-glucoside | 31.79 ± 1.36 b | 35.79 ± 0.64 a | 35.20 ± 0.75 a |
| Quercetin-3-O-glucuronide | 0.75 ± 0.04 b | 0.95 ± 0.05 a | 0.74 ± 0.05 b |
| Kaempferol-3-O-glucoside | 2.83 ± 0.12 a | 3.03 ± 0.06 a | 2.86 ± 0.11 a |
| Kaempferol-3-O-glucuronide | 1.99 ± 0.02 b | 2.19 ± 0.03 a | 2.11 ± 0.07 a |
| Isorhamnetin-3-O-glucoside | 0.18 ± 0.01 b | 0.22 ± 0.02 a | 0.20 ± 0.02 ab |
| Ʃ Flavonols | 43.00 ± 1.35 b | 48.51 ± 0.63 a | 47.87 ± 1.28 a |
| EGCG | 9.09 ± 0.33 a | 9.87 ± 0.36 a | 9.88 ± 0.30 a |
| ECG | 13.23 ± 0.18 b | 16.02 ± 0.24 a | 13.57 ± 0.20 b |
| Gallocatechin | 59.78 ± 1.19 b | 63.62 ± 2.48 ab | 65.71 ± 0.93 a |
| Epigallocatechin | 14.17 ± 0.15 b | 13.60 ± 0.15 c | 15.25 ± 0.28 a |
| Catechin | 25.12 ± 0.56 b | 30.35 ± 1.24 a | 29.24 ± 0.67 a |
| Epicatechin | 10.56 ± 0.30 b | 12.87 ± 0.07 a | 13.21 ± 0.56 a |
| PB1 | 61.21 ± 2.46 c | 98.34 ± 2.31 b | 112.76 ± 5.00 a |
| PB2 | 5.58 ± 0.10 b | 6.79 ± 0.49 a | 6.54 ± 0.17 a |
| PB3 | 2.48 ± 0.01 c | 2.94 ± 0.07 b | 3.29 ± 0.10 a |
| PB4 | 8.60 ± 0.16 c | 10.52 ± 0.56 b | 11.96 ± 0.01 a |
| PA1 | 1.69 ± 0.03 b | 1.94 ± 0.07 a | 1.73 ± 0.05 b |
| Ʃ Flavan3-ols | 211.52 ± 3.53 c | 266.87 ± 3.93 b | 283.15 ± 5.44 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mucalo, A.; Budić-Leto, I.; Zdunić, G. Effect of Sequential Fermentation with Lachancea thermotolerans/S. cerevisiae on Aromatic and Flavonoid Profiles of Plavac Mali Wine. Foods 2023, 12, 1912. https://doi.org/10.3390/foods12091912
Mucalo A, Budić-Leto I, Zdunić G. Effect of Sequential Fermentation with Lachancea thermotolerans/S. cerevisiae on Aromatic and Flavonoid Profiles of Plavac Mali Wine. Foods. 2023; 12(9):1912. https://doi.org/10.3390/foods12091912
Chicago/Turabian StyleMucalo, Ana, Irena Budić-Leto, and Goran Zdunić. 2023. "Effect of Sequential Fermentation with Lachancea thermotolerans/S. cerevisiae on Aromatic and Flavonoid Profiles of Plavac Mali Wine" Foods 12, no. 9: 1912. https://doi.org/10.3390/foods12091912
APA StyleMucalo, A., Budić-Leto, I., & Zdunić, G. (2023). Effect of Sequential Fermentation with Lachancea thermotolerans/S. cerevisiae on Aromatic and Flavonoid Profiles of Plavac Mali Wine. Foods, 12(9), 1912. https://doi.org/10.3390/foods12091912