3.2. Molecular Genetics
The amplification with at least one primer pair was successful for twelve (out of 20) individuals from four different samples (PA13, PA21, PA22, PA24). After the lower-quality sequences were discarded, the twelve sequences for COI, seven for 28S, and ten for 18S were obtained (
Table 2). No internal stop codons were found in COI sequences.
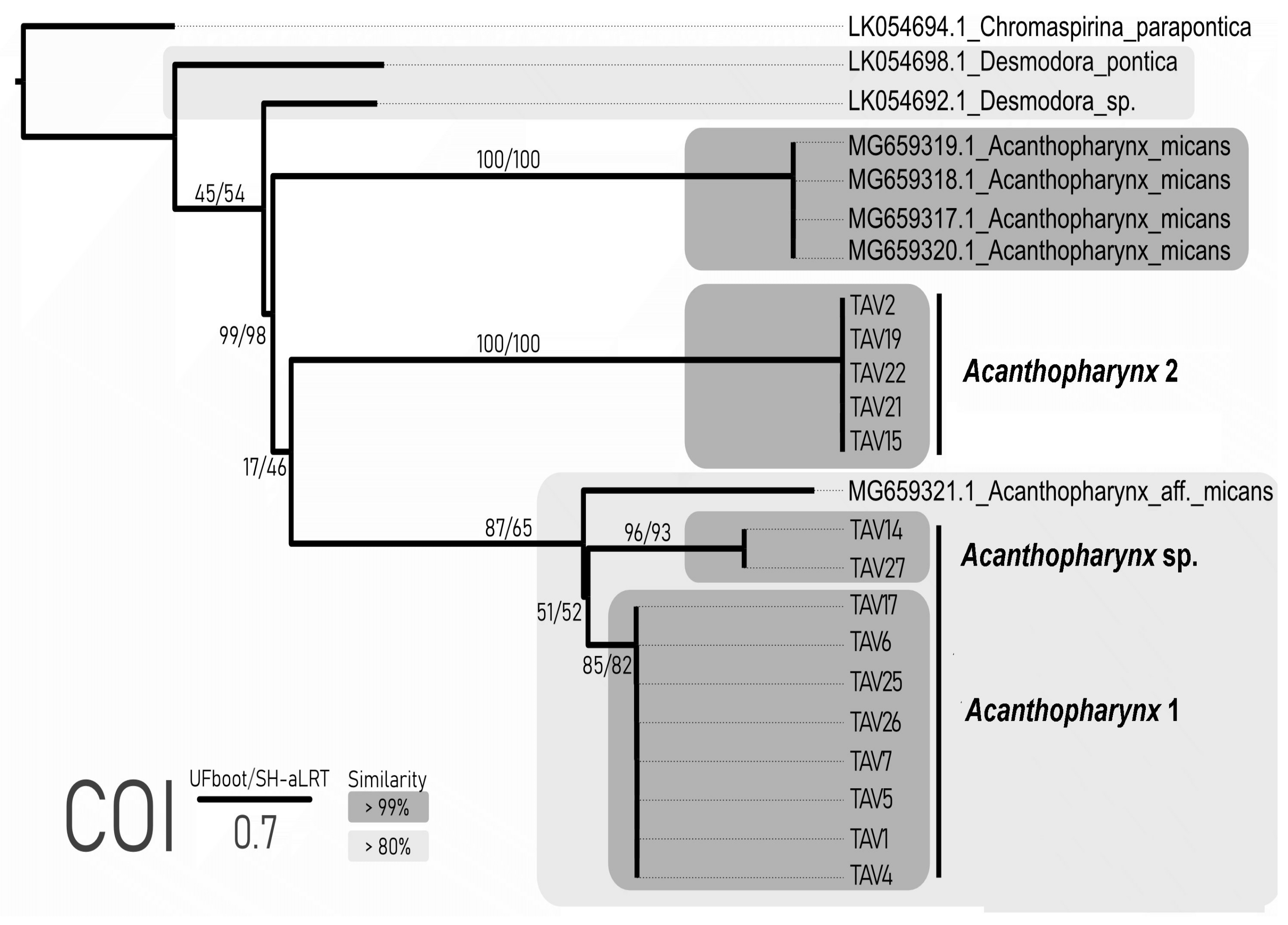
After primer and poorly read-end clipping, the length of the COI sequences was 372 bp. The number of differences in COI within the Acanthopharynx 1 was 0–52 bp (100–86% of identity). For Acanthopharynx 2, it spans 0–2 bp (100–99% of identity). Thus, the Acanthopharynx 1 comprises two groups of sequences. The number of substitutions within each sequence was equal to or less than 2 bp, with less similarity between the groups (86%, 52 substitutions).
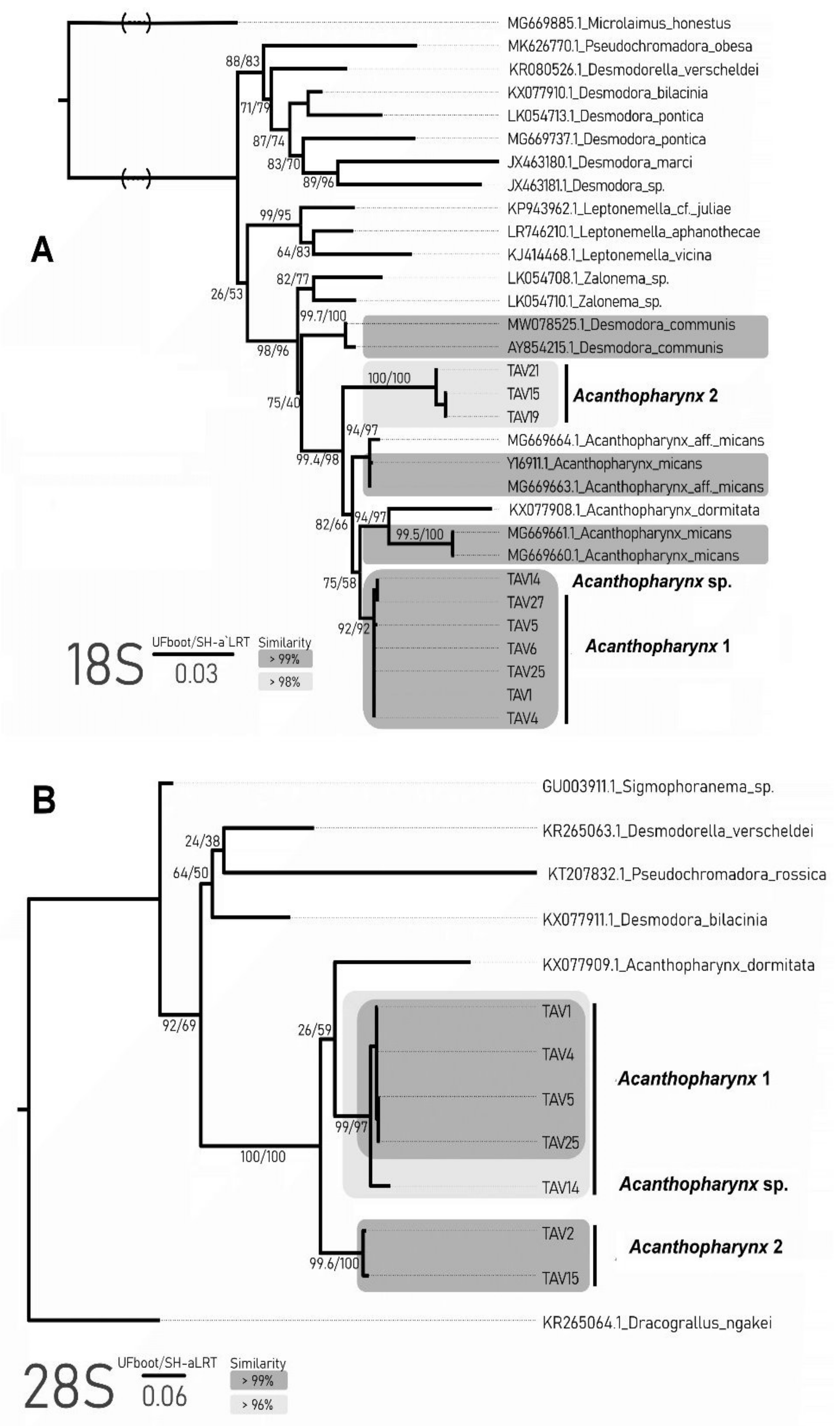
The COI alignment (including Genbank data) comprised 24 sequences and 372 sites, of which 181 were variable (160 parsimony informative). The sequences of
Acanthopharynx 1 and
Acanthopharynx 2 formed three clades sister to
Acanthopharynx micans and
Acanthopharynx aff.
micans representatives from Genbank with high to moderate UFboot/ SH-aLRT values (
Figure 5). The clade of
Acanthopharynx 1 formed two distinct clades, which is well matched with the similarity data (86% similarity between mentioned clades). The ABGD analysis with both distance metrics showed that an evident barcoding gap exist and all three mentioned clades of the
Acanthopharynx are genetically distinct. The mPTP method based on our phylogenetic tree recovered all three clades of the
Acanthopharynx as independent evolutionary entities (
p = 0.001).
The lengths of the 18S sequences obtained after primer clipping varied within 822–894 bp for
Acanthopharynx 1 and 847–863 bp for
Acanthopharynx 2. All sequences of
Acanthopharynx 1 were highly similar (99.6–100%). The similarity within the second species comprised 98.7–98.9%. The similarity between both morphospecies was less than 96%. The alignment of the 18S locus used for phylogeny reconstruction included 31 sequences with a length of 799 bp, of which 246 were variable and 138 were parsimony informative. All
Acanthopharynx species are grouped into one highly supported clade. The phylogenetic relationships of the other genera are not clear. Both morphospecies formed distinct clades sister to other
Acanthopharynx species (
Figure 6A). The results of ABGD and mPTP analysis also support the distinctiveness of these two clades.
The obtained sequences for 28S were shorter than 18S and comprised 666 to 692 bp for
Acanthopharynx 1. The length of
Acanthopharynx 2 sequences was 669 bp and 667 bp, with 98.1% similarity between them. The similarity within the first species varied from 96.1% to 99.9%, which is explained by the higher dissimilarity of one sample (TAV14). Without this individual, the similarities between other specimens were more than 99%. The 28S alignment was 655 bp long, with 332 variable sites, of which 200 were parsimony informative. The phylogenetic reconstruction showed that
Acanthopharynx species formed one clade sister to other genera (
Figure 6B). Two well-supported clades corresponded to morphospecies
Acanthopharynx 1 and 2. The third one (TAV14) belongs to the
Acanthopharynx 1 clade but is distinguished from its other representatives. The ABGD and mPTP analyses support the presence of three clades of
Acanthopharynx apart from the Genbank data.
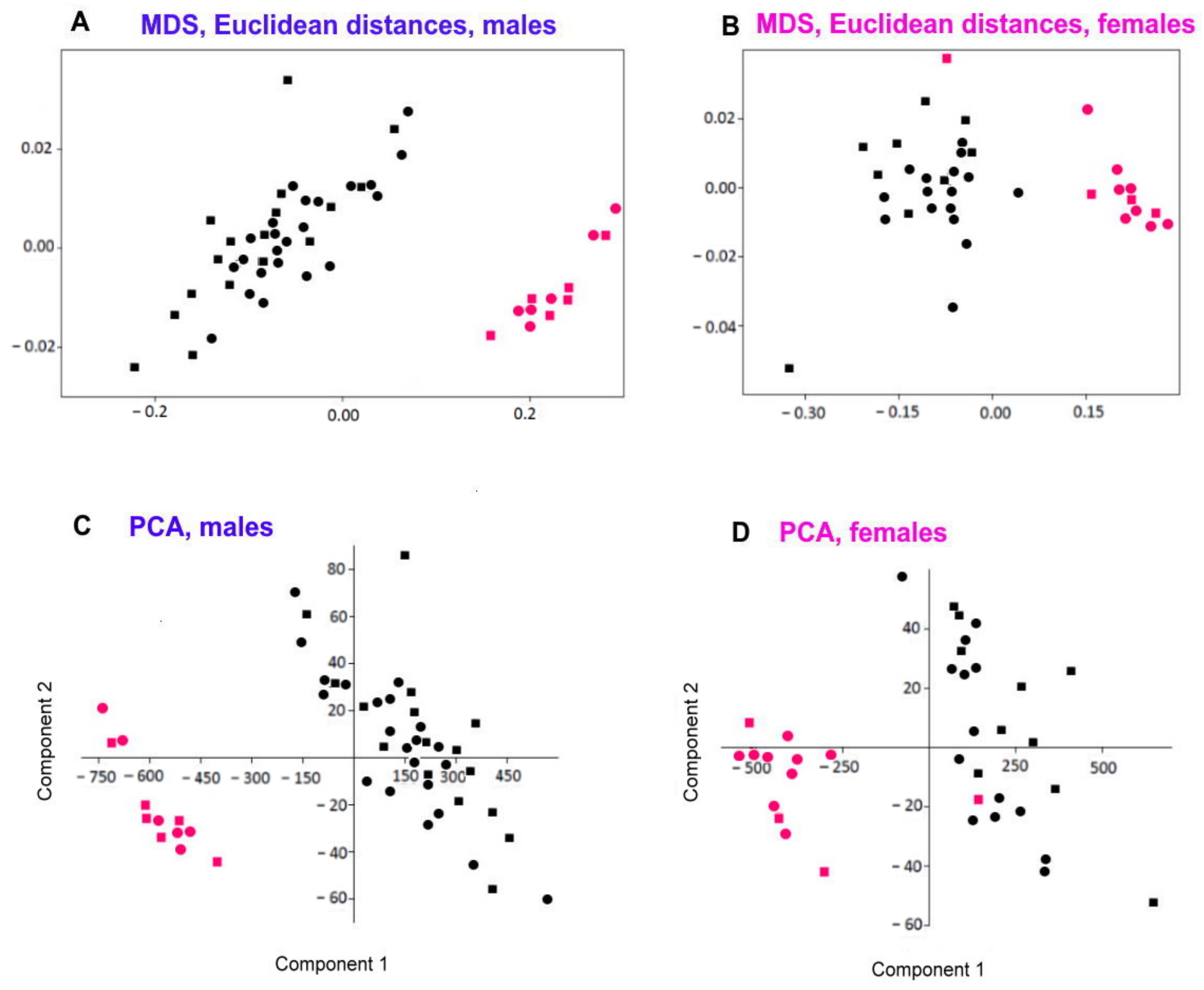
According to the species delimitation and phylogenetic tree analysis based on three loci (18S, 28S, COI), the studied specimens formed at least two distinct clades: Acanthopharynx 1 and 2. Two loci (28S, COI) support the existence of the third clade within the Acanthopharynx 1: specimen TAV14 and TAV27 as a distinct evolutionary entity. These two are probably cryptic species.
According to the molecular analyses, the Acanthopharynx 1 species fits into the same clade with specimens designated as Acanthopharynx aff. micans (COI) or with specimens designated as Acanthopharynx micans, Acanthopharynx aff. micans and Acanthopharynx dormitata (18S). Since Acanthopharynx 1 differs distinctly from A. dormitata in structural characters and morphometrics and at the same time does not differ or scarcely differ from descriptions of A. micans (see below in more detail), we recognize Acanthopharynx 1 as Acanthopharynx micans.
Acanthopharynx 2 is clearly separated from Acanthopharynx 1 in all three loci (COI, 18S, 28S). Since Acanthopharynx 2 also differs from Acanthopharynx 1 and other Acanthopharynx species by the shape of the pharynx and some morphometric characters, we consider it a new species for science and designate it as Acanthopharynx parvus sp. n. (see taxonomic part below).
As for individuals TAV14 and TAV27, prior to DNA extraction, they were examined under a light microscope and designated as Acantopharynx 1. Since no particular features in their morphology and morphometry were revealed, the status of these two specimens remains unresolved.
3.3. Taxonomy
Acanthopharynx is one of the earliest established genera of free-living marine nematodes [
26]. Since then, eighteen nominal species have been discovered worldwide [
27,
28]—yet some of those species were later considered junior synonyms or referred to other genera. Genus
Acanthopharynx can be rather easily recognized from other genera of Desmodoridae owing to numerous apical (cephalic and subcephalic) setae on the head and peculiar pharynx shape. Instead, identifying species may be difficult because the species differ from each other in fine details, such as supplementary papillae and pores, which are often missing in earlier descriptions. Some species described before the latter half of the twentieth century are treated as
species inquirendae, which means their validity may be restored after a thorough redescription based on specimens from the type locality.
The descriptions of species issued later, starting from
Acanthopharynx denticulata Wieser, 1954, are characterized by a high level of detail and allow us to understand the morphology of the
Acanthopharynx [
8,
29,
30,
31]. Thus, Leduc & Zhao [
8] constructed a schematic pattern of sensilla on the head capsule based on SEM observation; they also provided molecular sequences (SSU and D2-D3 of LSU) of
A. dormitata and showed that
Acanthopharynx forms a basal clade to the Desmodorinae/Spiriniinae. Though Wieser [
29] provided a dichotomous key for the identification of eight species known at that time, the lack of necessary details and hence the nonuniformity of morphological data between formerly and later described species encumber the development of an upgraded species identification key.
Order Desmodorida de Coninck, 1965
Family Desmodoridae Filipjev, 1922
Subfamily Desmodorinae Micoletzky, 1922
Genus Acanthopharynx Marion, 1870
(=Xanthodora Cobb, 1920; Brachydesmodora Allgén, 1932)
Diagnosis
amended after Tchesunov 2014 and Leduc & Zhao 2016 [
8,
32].
Desmodoridae, Desmodorinae. Cuticle distinctly annulated between the cephalic capsule and terminal tail cone; no lateral differentiation. Cephalic capsule non-articulated, smooth. Labial region is bordered by the cephalic capsule by a fine suture. The anterior sensilla is concentrated close to the apex and is composed of six inner labial papillae (may be indistinct), six outer labial papillae, and four cephalic setae; a number (about ten or twelve) of subcephalic setae installed in the same crown with cephalic setae; the subcephalic setae are nearly equal in length and shape to cephalic setae and hardly distinguishable from them. Minute somatic setae arranged in irregular rows along the body. Amphideal fovea spirally coiled in one turn, round or rarely longitudinally oval in outlook; the cuticle spot in the center of the fovea differs from the cuticle outside the fovea. Buccal cavity armed with a solid movable dorsal tooth and two transversal ventrolateral rows of minute denticles; additional denticles may be present. The pharynx is muscular and has a thick internal cuticular lining along its entire length; the posterior 40–60% part is either sharply swollen as an elongated bulb or gradually swollen to its posterior end. Precloacal midventral supplementary organs usually present as a series of tiny papillae or pores and larger papilla close to the cloaca. Spicules arcuate, slightly knobbed. Gubernaculum as a short bar contiguous to spiculum. Tail short, conical, slightly bent ventrally, with a smooth terminal cone.
Type species Acanthopharynx affinis Marion, 1870.
Annotated species list (valid species names in bold)
- (1)
Acanthopharynx affinis Marion, 1870. Marion, 1870a: 36–37, Plate K, Figure 4–4b [
26]; Mediterranean. Schuurmans Stekhoven 1942:243–244, Figure 13A–C [
33]; Mediterranean.
- (2)
Acanthopharynx brachycapitata (Allgén, 1947). Allgén, 1947: 148–149, Figure 51a–c [
34] (only female) (as
Desmodora brachycapitata); Gulf of Panama. Gerlach 1963: 77 [
35] (transfer to
Acanthopharynx). Verschelde et al. 1998:82 [
36] (
species inquirenda).
- (3)
Acanthopharynx denticulata Wieser, 1954. Wieser, 1954a: 36, Figure 113a–d [
29] (males, females, juveniles); North Chile, littoral algae, sheltered. Armenteros et al. 2014: 5–8, Figures 1A–D and 2, Table 1 [
31] (males, females, juveniles); Cuba, south coast, 2 m, sand flat (lapsus
denticulatus). Cuban males differ from Chilean males in some measurements such as body length (1234–2780 versus 2170–2780 μm), index b (5–7 versus 8.1–9.7), index c (16–21 versus 22–31)—however, Armenteros et al. [
31] consider the differences as intraspecific.
- (4)
Acanthopharynx distechei Decraemer & Coomans, 1978. Decraemer & Coomans, 1978:515–519, Figure 2A–E [
30] (male, female, juveniles); Great Barrier Reef, Lizard Isl., mangrove swamp.
- (5)
Acanthopharynx dormitator Leduc & Zhao, 2016. Leduc & Zhao, 2016: 908–916, Figures 1–6 [
8] (males, females); New Zealand, Wellington, lower intertidal zone, red seaweed partially covered in sediments on boulders.
- (6)
Acanthopharynx japonica Steiner & Hoeppli, 1926. Steiner & Hoeppli, 1926: 551–555, 568–569, Figures A–F [
37]; Pacific coast of Japan.
- (7)
Acanthopharynx merostomacha (Steiner, 1921). Steiner, 1921: 52–54, Tafel 3, Figure 12a–c [
38] (one juvenile) (as
Desmodora merostomacha); Red Sea. Schuurmans Stekhoven 1943: 363 [
39] (transfer to
Acanthopharynx). Allgén 1951: 300–301, 389 [
40] (male, female); Pacific (Honolulu, California, Bay of Panama). Allgén, 1959: 114 [
41]; Falkland Islands, South Georgia. Verschelde et al., 1998: 82 [
36] (
species inquirenda).
- (8)
Acanthopharynx micans (Eberth, 1863) Marion, 1870. Eberth, 1863: 4, 6, 12, 28, Table 1, Figures 1–5 [
42] (male, female) (
Odontobius micans); Mediterranean, Nizza. Marion 1870: 6 [
26] (transfer to
Acanthopharynx). Micoletzky, 1924: 148–151, Figure 1a–b [
43]; Mediterranean, Red Sea. Schuurmans Stekhoven 1942: 244 [
33] (
Acanthopharynx marioni). Wieser 1954: 199, Abbildung 13 [
44] (males); Mediterranean. Gerlach 1963: 93–94, Table 9, Figure g–k [
35] (male); Maldives. Schuurmans Stekhoven 1950: 121, Figure 71A–C [
45] (male) (as
Acanthopharynx seticauda); Mediterranean.
- (9)
Acanthopjharynx micramphis Schuurmans Stekhoven, 1942. Schuurmans Stekhoven, 1942: 245–247, Figure 14A–C [
33] (male, juveniles); Mediterranean, Ibiza. Despite presence of a male in the type series, all the body dimensions are given for a juvenile. Nonetheless, the males of the species can be recognized owing to the small amphideal fovea (14% cbd) and short conical tail (c’ 1.75).
- (10)
Acanthopharynx nuda (Cobb, 1920) Gerlach, 1963. Cobb, 1920: 317–318, Figure 98 [
46] (male, female) (as
Xanthodora nuda); Indonesia, Larat Island. Gerlach 1963: 94 [
35] accepted
Xanthodora as a synonym of
Acanthopharynx.
- (11)
Acanthopharynx parva sp. n. Present paper.
- (12)
Acanthopharynx perarmata Marion, 1870. Marion, 1870: 34–35, Plate K, Figure 1–1f [
26] (female); Mediterranean. Schuurmans Stekhoven 1942: 245 [
33]; Mediterranean, Naples. Schuurmans Stekhoven 1950: 118–120, Figure 69A–C [
45] (male); Mediterranean, Villefranche. Despite the incompleteness of the original and subsequent descriptions, the species can be recognized by a very short tail and three prominent wartlike papillae (orig. “excrescences”) on the ventral side of the tail.
- (13)
Acanthopharynx rigida Schuurmans Stekhoven, 1950. Schuurmans Stekhoven, 1950: 120–121, Figure 70A–C ([
45] male); Mediterranean, Villefranche.
- (14)
Acanthopharynx setosissima Schuurmans Stekhoven, 1943. Schuurmans Stekhoven, 1943: 362–363, Figure 31A–B [
39] (female); Mediterranean, Alexandria. The original description based on a single female is incomplete and does not allow for the exact identification of this species. Hence, we consider this species as a
taxon inquirendum. - (15)
Acanthopharynx similis (Allgén, 1932) Gerlach, 1963. Allgén, 1932: 463–464, Figure 23a–b [
47] (only juvenile) (
Desmodora (
Brachydesmodora)
similis); Campbell Island, slime of cyanophycean algae. (=Allgén 1932: 133–135, Figure 21a–c [
48]); Gerlach 1963: 77 [
35] (transfer to
Acanthopharynx). Verschelde et al. 1998: 82 [
36] (
species inquirenda).
Acanthopharynx micans (Eberth, 1863) Marion, 1870.
Material Examined
Forty-one males and twenty-eight females in permanent glycerin slides have been observed, measured, partly pictured, and drawn. Some slides (no catalog numbers) are deposited in the nematode collection of the Center of Parasitology, A.N. Severtsov Institute of Ecology and Evolution of the Russian Academy of Sciences, Moscow, Russia.
Locality
South coast of Cuba in the vicinity of Cienfuegos city, Ancόn Beach, 21°71′01.53″–21°75′31.79″ N and 79°99′39.96″–80°02′75.64″ W, depths 9–16 m, sponges, 17–18 November 2019.
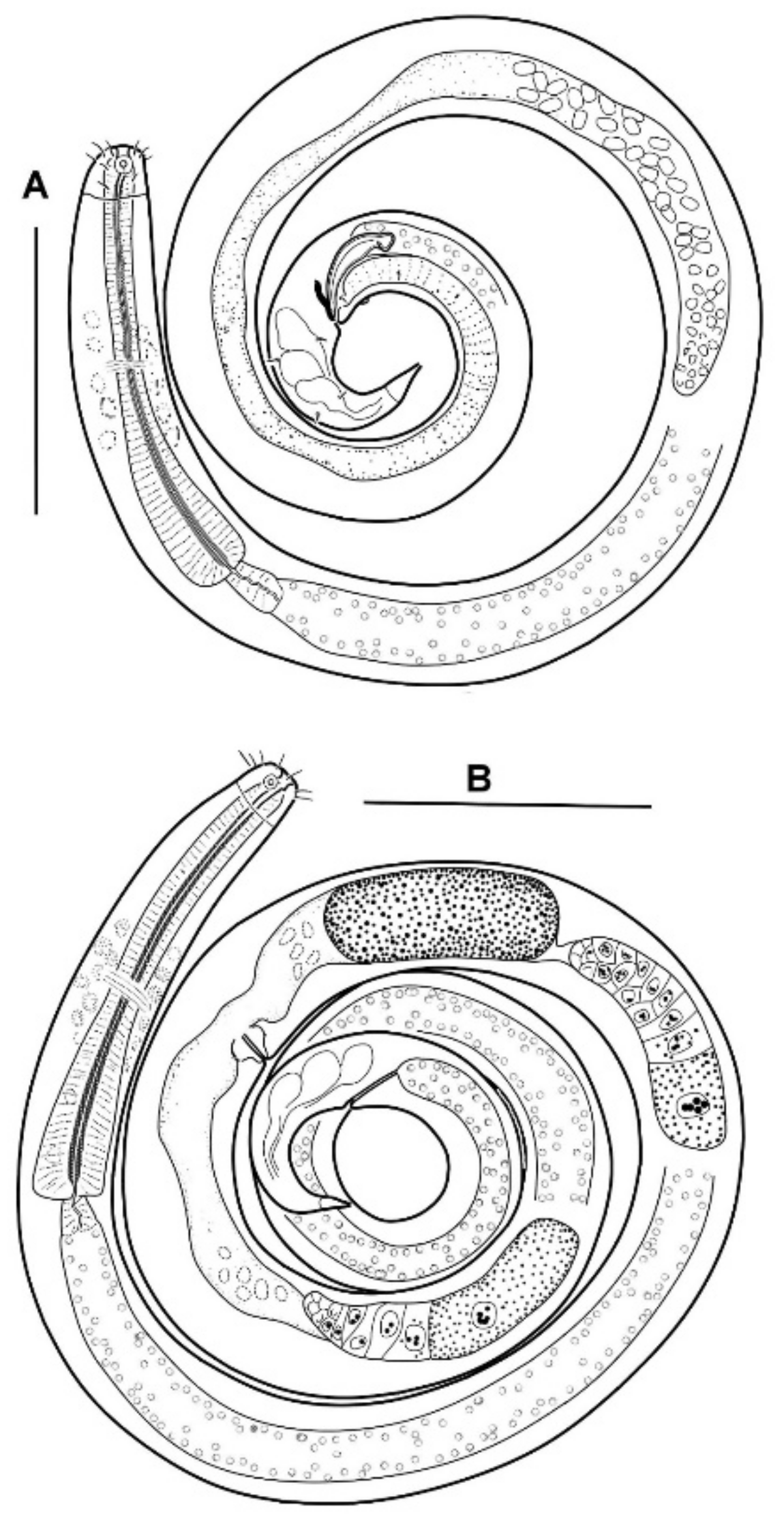
Description
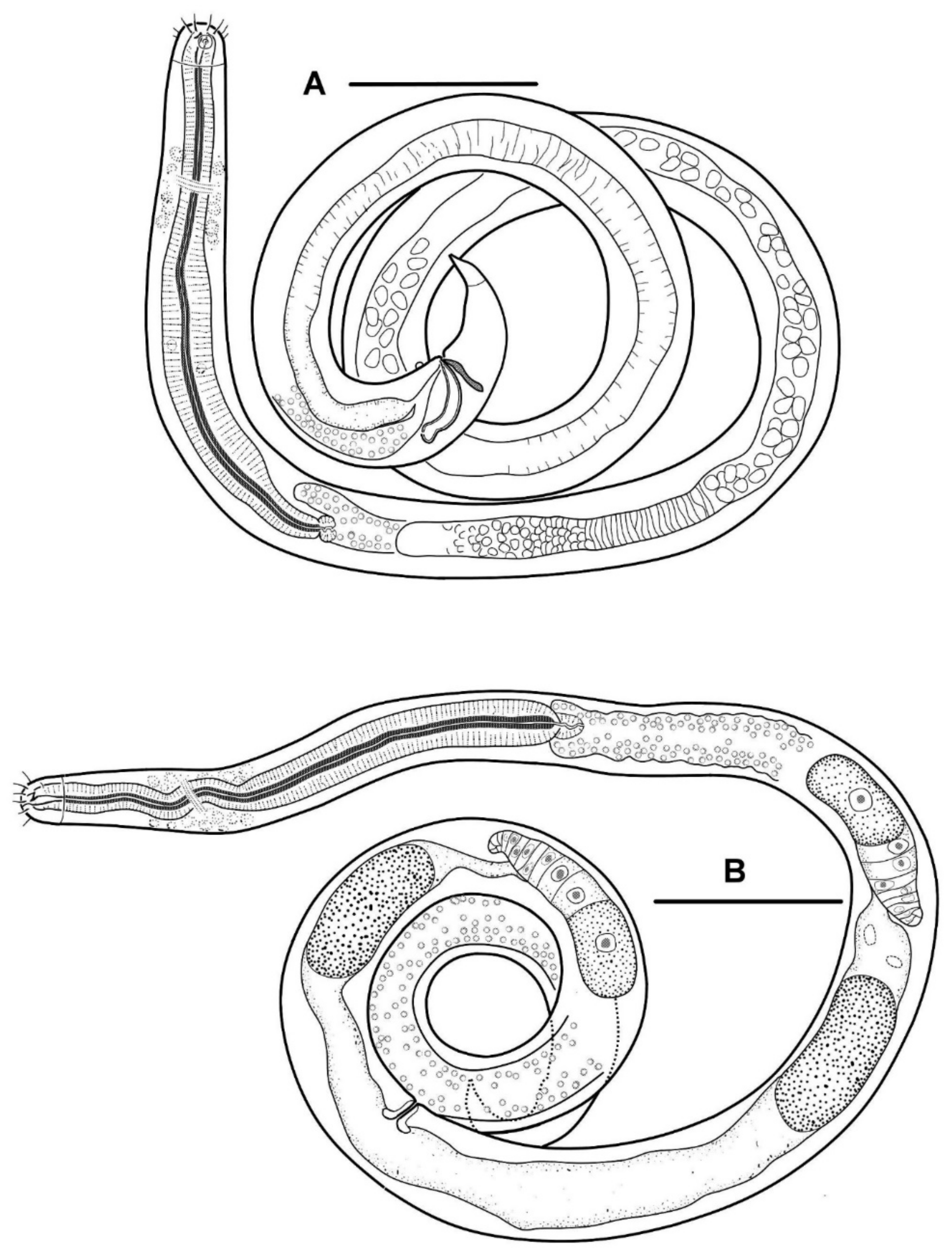
Body slender, cylindrical. The cuticle is finely but distinctly annulated; the annulation is uniform along the entire body from the cephalic capsule to the tail terminal cone. In a male, there are 13 annules per 20 μm just posterior to the cephalic capsule, 21–22 annules per 20 μm in the midbody, 20 annules per 20 μm just anterior to the cloaca, 21 annules per 20 μm just anterior to the tail terminal cone.
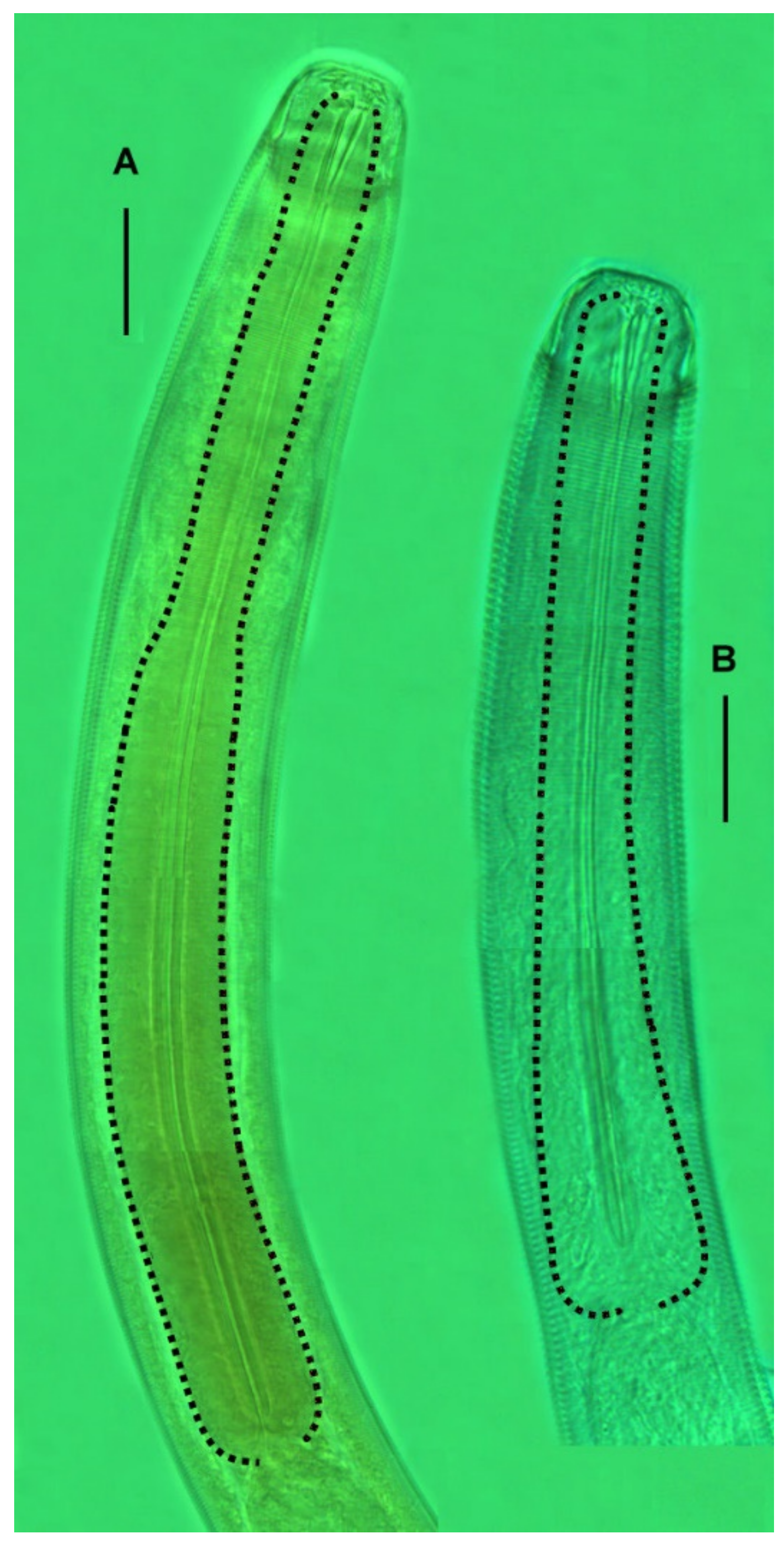
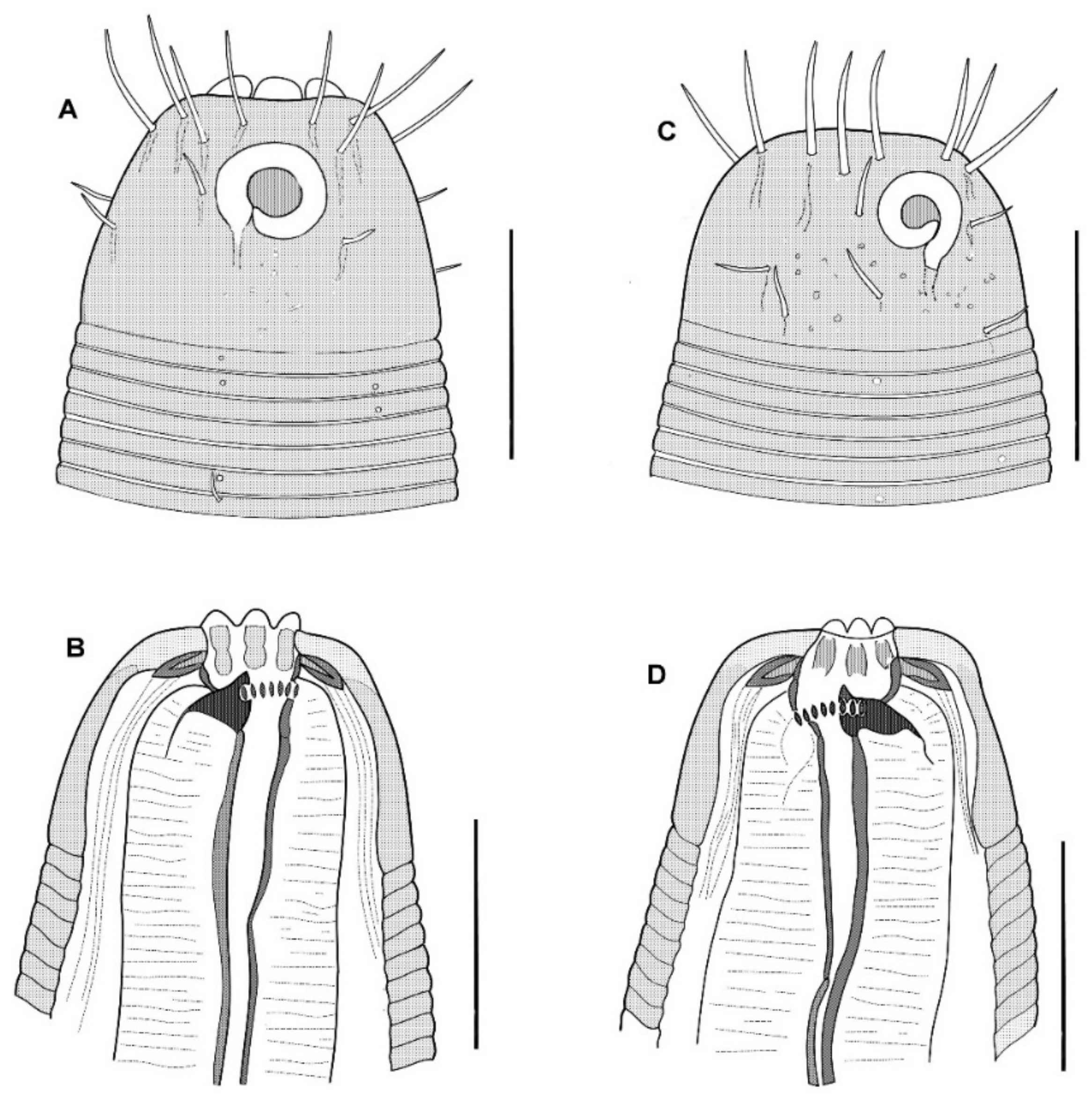
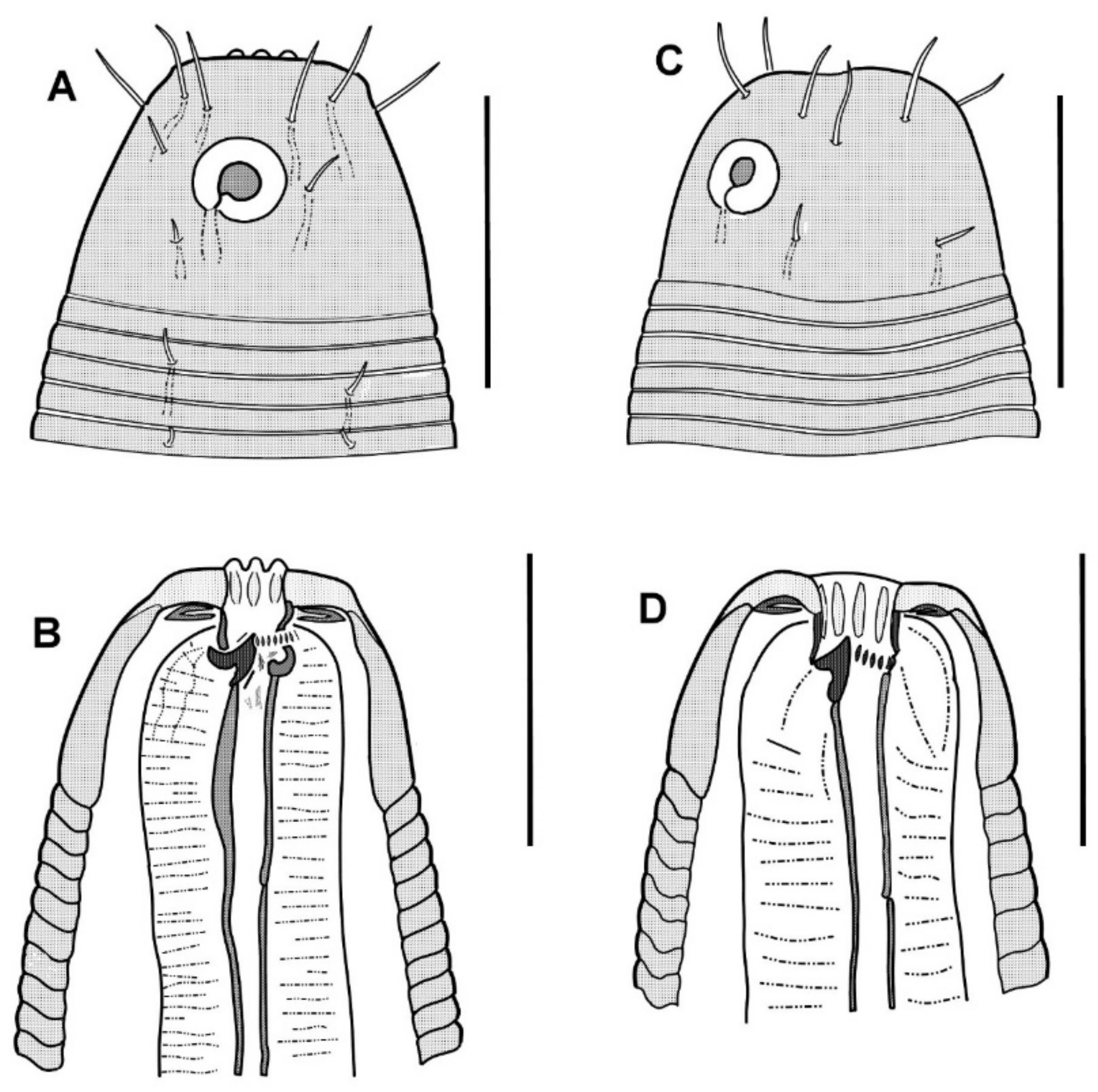
The cephalic capsule is shaped as a rounded truncate cone. The cuticle of the cephalic capsule is smooth. The labial region is distinctly bordered by a circular furrow. The mouth opening is surrounded by six small lips, which are raised up in some of the specimens. No sensilla were observed in the labial region. There are six papillae (evidently, outer labial sensilla) just posterior to the labial region. For all of the setae, altogether up to 16 or 22 in number, located apically and subapically on the head, are all nearly equal in length and directed forward. The cephalic setae in the lateromedial position are distinguished from other apical setae neither by length nor width. There are two additional sublateral setae between the cephalic setae on either lateral side of the head. In addition to the apical and subapical setae, other more posterior setae on the cephalic capsule are much smaller and sparse, and they are arranged in about ten loose longitudinal rows. The rows of short setae or papillae continue posteriad along the body. Amphideal fovea medium-sized, situated anteriorly on the cephalic capsule, spiral in one turn with a central spot, round in outline.
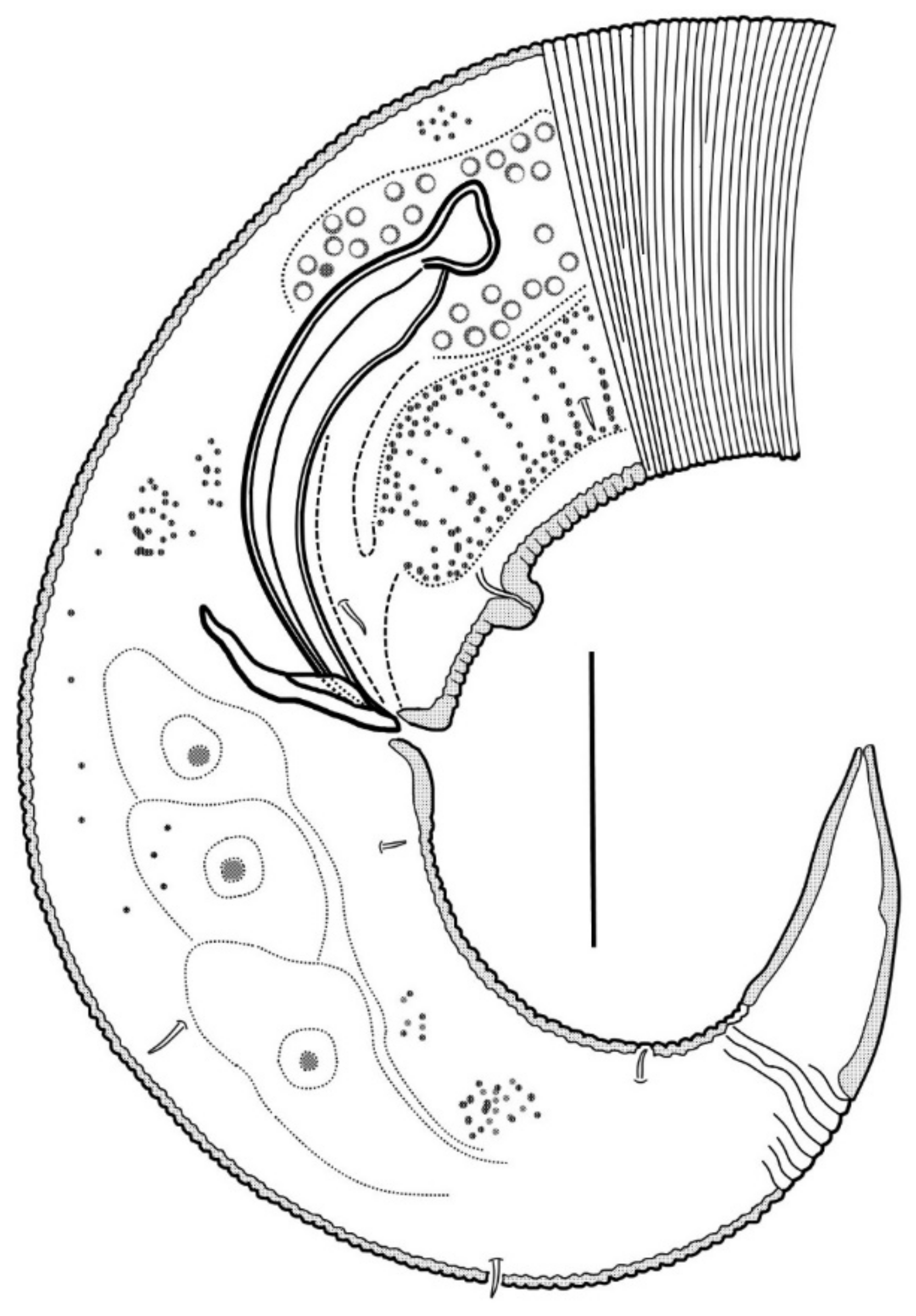
In the optical section, the somatic cuticle around the cheilostoma light homogeneous is distinctly separated from the main part of the cephalic capsule. The walls of the cheilostoma are complicated with short longitudinal rugae, evidently, there are twelve in number. The lower layer of the apical cheilostomatal cuticle around the mouth differentiated into cuneiform light-refracted structures. The cuticle of the cephalic capsule looks dense and non-vacuolated, which is thickened considerably to the posterior edge. Pharyngostoma consists of two parts, anterior about cup-shaped gymnostoma and elongated tightly conoid stegostoma, both with cuticularized walls. There is a prominent solid fang-shaped dorsal tooth at the anterior edge of stegostoma. Two lateroventral comb-like rows of minute denticles are opposed to the dorsal tooth. Both dorsal teeth and rows of denticles can push out a bit. The denticles, however, are very fine and hardly discernible in many specimens. The entire stegostoma are elongated and narrowing posteriad is evident, and its posterior end is marked with a notch. The pharynx is evenly muscular throughout its length, with a very distinct thick internal cuticular lining. The posterior portion over half of the entire pharynx constitutes a long bulb-like widening, which is distinctly separated from the narrow anterior part; the internal cuticular lining is also enlarged in the posterior widening of the pharynx. A hardly visible nerve ring is located at the posterior end of the narrow anterior part of the pharynx. The cardia is obscure.
No ventral gland is visible.
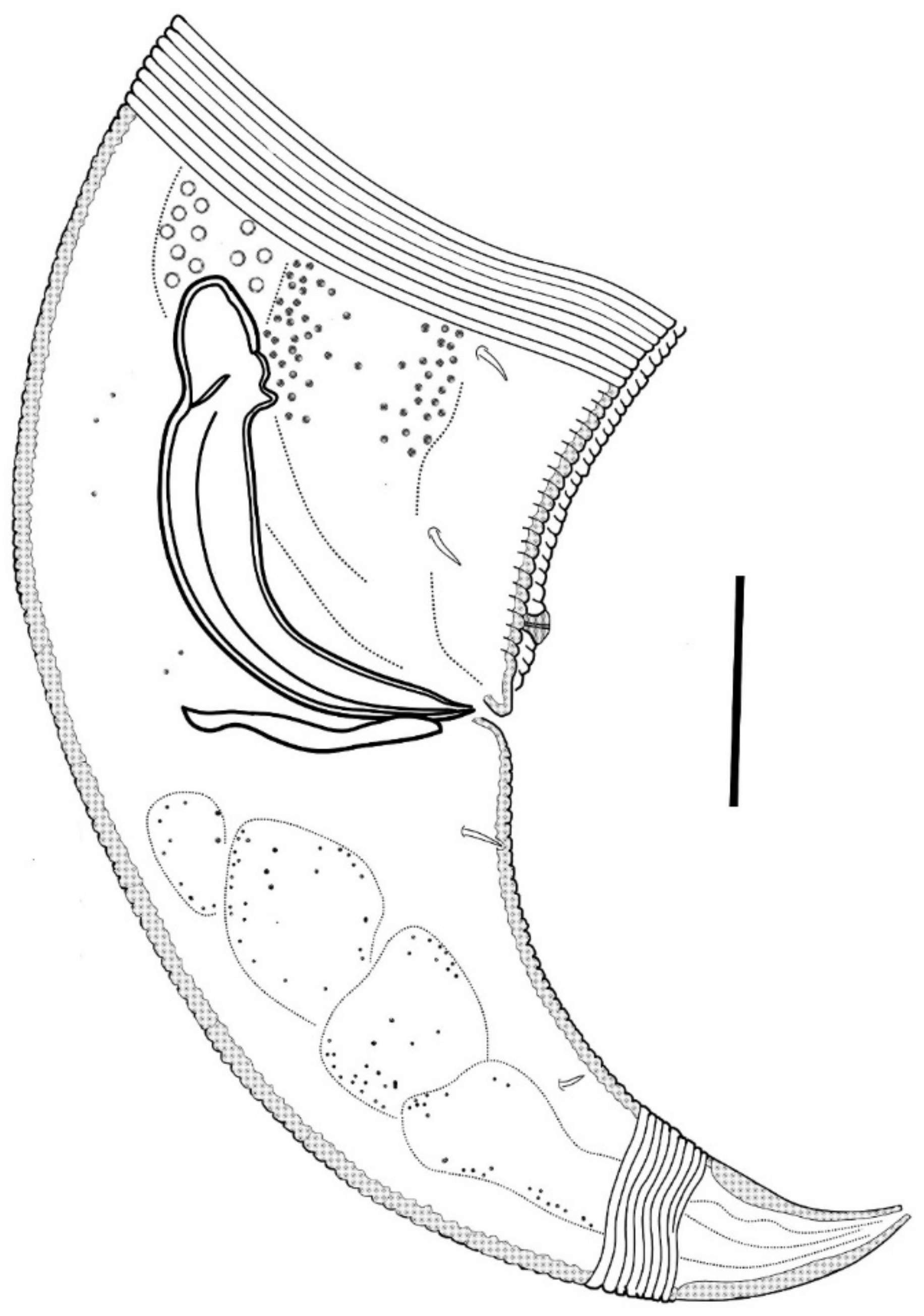
The male reproductive system is monorchic, and the testis are situated to the right of the intestine. The spermatozoa are irregularly ovoid, with granular content. Spicules paired, equal, arcuate, with anterior moderately differentiated proximal knobs and pointed distal tips. The gubernaculum consists of paired stick-like bars perpendicular to the longitudinal body axis. There is a long midventral precloacal shallow furrow extending anteriad from the cloaca ridge. A low midventral ridge rising from the furrow bears a row of hardly discernible supplementary pores and a posteriormost supplementary organ with the appearance of a soft wart close to the cloaca.
Tail conical, pointed, slightly curved ventrally, with few small pre- and postanal subventral setae. Terminal cone with smooth cuticle. No sensilla was found on the tail and in the precloacal area.
Remarks
Acanthopharynx micans is apparently the most common and widespread species of the genus: formerly,
A. micans was recorded in several localities of the Mediterranean, Red Sea, and Maldive Islands [
27]. The species was found only on shallows and not so much in sediments as on corals, algae, and mussel banks [
33,
35,
45,
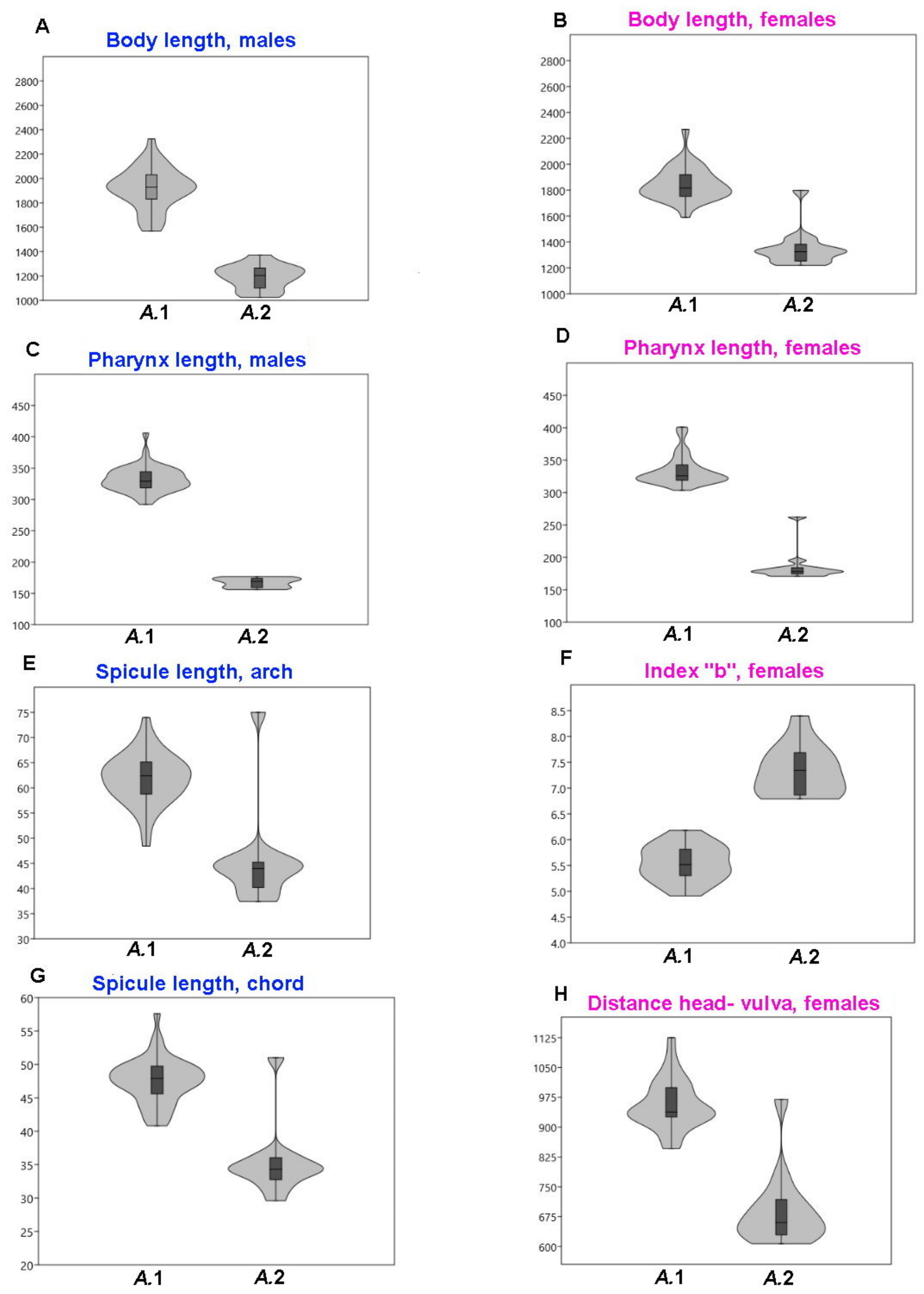
49]. The cuban specimens do not differ from those of other regions in structural details. However, they are distinguished by larger body size (
Table 6), which may relate to their habitation in sponges under the enhanced feeding condition.
Another
Acanthopharynx species previously known in nearly the same location is
A. denticulata [
31], but this species has been found in bottom sediments, not in sponges. Morphometrically, Cuban
A. micans conforms with the sympatric
A. denticulata population in body length and body ratios but differs by a longer pharynx (292–406 versus 214–286 μm), pharynx shape with ellipsoid versus cylindrical posterior widening and precloacal midventral row of one prominent posteriormost papilla with a number of supplementary pores versus 13–16 equal supplementary pits.
Acanthopharynx parva sp. n.
Etymology
The species name (from the Latin “parvus,” little) reflects the smaller body sizes than the cohabiting Acanthopharynx micans.
Material Examined
All types of specimens are mounted in permanent glycerin slides. Holotype male (slide 50/12) and allotype female (slide 50/13) were deposited in the nematode collection of the Center of Parasitology, A.N. Severtsov Institute of Ecology and Evolution of the Russian Academy of Sciences, Moscow, Russia. Twelve male and thirteen female paratypes are deposited in the same collection.
Type Locality
The south coast of Cuba in the vicinity of Cienfuegos city, Ancόn Beach, 21°71′01.53″–21°75′31.79″ N and 79°99′39.96″–80°02′75.64″ W, depths 9–16 m, sponges, 17–18 November 2019.
Description
The body is cylindrical with a rounded head end and a short conical tail. The cuticle is faint but distinctly annulated except for the cephalic helmet (cephalic capsule) and terminal tail cone. In the holotype male, there are 13 annules within 20 µm just behind the cephalic capsule, 25 annules within 20 µm just behind the cardia, 30 annules within 20 µm in the midbody, 22 annules within 20 µm in the midtail (dorsal convex side). In a paratype female, there are 12 annules within 20 μm just behind the cephalic capsule, and 20 annules within 20 μm in the midbody.
The cephalic capsule, which is non-annulated, is formed by a dense somatic cuticle. The cuticle of the cephalic capsule is light-refracted, non-sculptured, and non-vesiculated, having a thickened posteriad. There are about twelve short setae forming a subapical crown; four cephalic setae do not differ in size from other setae of the crown; the cephalic setae are identified based on their lateroventral position. Posterior to the subapical crown, the other setae on the cephalic capsule are minute and arranged irregularly posterior to the subapical crown. The transcuticular nerve canals of setae and amphid are very distinctly visible. There are sparse rows of tiny somatic setae extended along the body.
The amphideal fovea presents a circular interrupted ring with a cuticular spot in the center; actually, the fovea is a spiral coiled ventrally in one turn.
The mouth opening is surrounded by six small lips. The cheilostoma features levigate longitudinal rugae. The anterior part of the pharyngostoma features cuticularized walls, teeth, and denticles. The dorsal tooth is solid, claw-like, and movable; it is opposed by a pair, left and right lateroventral flanges of tiny denticles, also protrusible. The posterior part of the pharyngostoma is elongated and tight; its posterior ending is marked by a light flexure. There are cuniform solid cuticular structures on the internal surface of the circumoral apical cuticle; they are connected with longitudinal muscles and likely serve by opening the mouth.
Pharynx with the finest transversal muscular striation throughout its length. The posterior fourth to third of the pharynx is enlarged and in the shape of an elongated isosceles triangle. The internal cuticular lining of the pharynx is widened, especially in the posterior half. The cardia is external and narrow.
The male reproductive system is monorchic and situated to the right of the intestine. The spermatozoa is ovoid and have a granular content; they are about 8 × 4 µm in size. The paired spicules are equal, arcuate, distally pointed, and proximally knobbed. The gubernaculum presents as paired bars oriented dorso-ventrally. An only evident precloacal supplementary papilla is situated midventrally close to the cloacal opening. Precloacal midventral papillae or pores are not visible.
In females, the ovaries are antidromously reflected, anterior left and posterior right to the midgut.
Tail short conical, with three poorly discernible caudal gland cell bodies within. Cuticle of the tail hind part non-annulated, thus shaping a terminal cone.
Diagnosis
Acanthopharynx. Body length 1020–1440 μm, a 25–42, b 5.8–8.4, c 10.6–17.3, c’ 3–4.5. Cephalic setae 4–7.5 μm long. The amphideal fovea are spirally coiled in one turn and circular in outline. The buccal armature consists of a movable dorsal tooth and two transversal lateroventral rows of minute denticles. The pharynx is short and continually widened to the cardia without a sharply defined bulb. The spicules are 29–75 μm long. The precloacal midventral organs are only present with papilla. There were no distinct postcloacal supplementary organs.
Differential diagnosis
Acanthopharynx parva sp. n. differs from all the other Acanthopharynx species (with possible exception of A. denticulata) due to the shape of the pharynx gradually widening to the cardia and lacking a conspicuous elongate swelling or bulb. In addition to that, A. parva clearly differs from those Acanthopharynx species characterized by unique prominent features. Thus, A. parva differs from A. affinis by the presence of the prominent precloacal papilla; from A. denticulata by the absence of subventral teeth in the stoma (A. denticulata possesses smaller subventral teeth in addition to a large dorsal tooth and two rows of minute denticles) and the presence of prominent precloacal papilla; from A. distechei by longer cephalic setae (4–7.5 μm versus 3–3.5 μm), round versus longitudinally oval amphideal fovea, short spicules 29–75 versus 117 µm, oviparity versus viviparity; from A. dormitata by a discrepant set of precloacal supplementary organs (one prominent posteriormost papilla versus nine precloacal and two postcloacal papillae); from A. micramphis by presence of the preanal papilla (no supplementary organs not observed on A. micramphis), from A. nuda by another set of supplementary organs (an only conspicuous precloacal papilla versus six precloacal papillae along a distance equal to the tail length); from A. perarmata also by supplementary organs (an only precloacal papilla versus three almost equidistant papillae).
The differences in
Acanthopharynx parva from
A. japonica and
A. rigida are somewhat less evident since those species were described seventy or more years ago. Therefore, some structures, foremost the buccal armature and the pattern of the supplementary papillae need to be specified in detail. Morphometric differences of
A. parva from
A. japonica,
A. micans,
A. rigida and other species are summarized in
Table 6.
In addition, it is necessary to consider the relations of
A. parva to two cohabitated
Acanthopharynx species populations.
A. parva differs from
A. micans (present paper) in the shape of the pharynx (gradually broadening to the posterior end versus distinctly separated elongate widening) and lesser body length (1020–1440 μm versus 1569–2325 μm), pharynx length (156–195 μm versus 292–406 μm), c of males (10–15 versus 16–25).
A. parva differs from sympatric
A. denticulata [
31] by pharynx shape (posterior widening ellipsoid in
A. denticulata), index c of males (10–15 versus 16–21), c’ of males (3.2–3.9 versus 1.6–2.2), singular prominent precloacal papilla versus 13–16 equal supplementary pits.






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}